Questo post partecipa all'edizione speciale del carnevale della chimica il quale nel mese si settembre si unifica nell'argomento trattato con il carnevale della fisica. Il tema è "
CERCANDO TRACCE DI VITA NELL'UNIVERSO".
Invito tutti i lettori che approderanno a questa pagina di fare una capatina sul sito dell'associazione
CHIMICARE e sul sito del
carnevale della chimica per maggiori informazioni.
L'idea di scoprire una nuova forma di vita extraterrestre per decenni ha entusiasmato e continua ad entusiasmare non solo astronomi e astrobiologi, ma anche il grande pubblico e perchè no solleticato anche la fantasia di molti bambini, soprattutto la mia quando ero piccolo, quante ore perse a giocare e a fantasticare, dopo aver visto i vari alien, predator e star wars, su quali pianeti sconosciuti arrivassero i personaggi, interpretati magistralmente dai miei giocattoli con la mia voce narrante, e sulle peripezie da dover affrontare per un salvo ritorno a casa!
L'idea di non essere soli nell'universo, di essere l'unico esempio di forma di vita nella galassia è per molti un qualcosa di statisticamente improbabile, soprattutto se pensiamo alle varie forme di vita esistenti sul nostro pianeta, in grado di esplicare magistralmente svariate funzioni metaboliche e replicative e alle strategie che permettono ad alcuni microrganismi di sopravvivere in ambienti che sarebbero inospitali per la maggior parte delle specie viventi!
Ma può una forma di vita qualora la trovassimo su altri pianeti essere definita veramente aliena? Cosa intendiamo con forme di vita nell'universo e quante sono le probabilità di scoprire nuove forme di vita? Ovviamente nessuno lo sa, ma c'è chi prova ad esaminare il problema ponendosi quesiti di vario tipo, tra questi vi è Gerald Joyce dello Scrips Research institute di La Jolla; California.
Lo studio, pubblicato sulla rivista Plos One biology delinea le caratteristiche che dovrebbe avere una forma di vita aliena per definirla tale.
In particolar modo dobbiamo chiederci se il tipo di vita che eventualmente potremmo trovare in qualche remoto pianeta rispecchiasse le regole biologiche a cui gli organismi sul nostro pianeta sono soggetti e come la vita su altri pianeti, cosi come lo è stato milioni e milioni di anni addietro sul nostro, abbia potuto svilupparsi.
Il professor Gerald Joyce, cerca di discutere i requisiti fondamentali per l'esistenza di nuove forme di vita. "La vita si auto-riproduce, trasmette le informazioni ereditarie alla sua progenie, e subisce l'evoluzione darwiniana basata sulla selezione naturale."
Nel saggio, che potete leggere al link presente a fine post, fa riferimento a queste informazioni ereditabili come "bit" (per la vita come noi la conosciamo, questo include le quattro basi del DNA), e ci spiega come, anche se l'evoluzione darwiniana rimescola e crea nuove combinazioni di tali informazioni, ciò non definisce una nuova forma di vita o la creazione di una specie "aliena".
Infatti, ad oggi nessuna forma di vita veramente nuova è stata scoperta - sia in ambienti estremi sulla Terra o su altri pianeti - che contenga nuove caratteristiche, differenti da quelle a cui sono soggetti gli organismi sul nostro pianeta.
Siamo abituati a studiare e a pensare la vita sostanzialmente come una cellula al cui interno vi è un genoma contenente informazioni genetiche sottoforma di nucleotidi, informazione che viene tradotta, concretizzata dalla cellula tramite la costruzione delle proteine.
Ne consegue che se vogliamo uscire fuori dallo schema nel quale ci inglobano le leggi della chimica e della biologia come noi le conosciamo, occorre, secondo joyce, ragionare in maniera diversa, chiedendoci soprattutto come e quali possano essere le condizioni che permettano il verificarsi di eventi che portino alla creazione di componenti molecolari in grado di creare una informazione che possa essere trasferita di generazione in generazione, che muti dando vita a diverse combinazioni e a nuova vita.
Un sistema chimico ed un sistema biologico.
Secondo l'autore la vita potrebbe nascere tramite due possibilità: 1) potrebbe derivare, direttamente o attraverso la chimica, o da un sistema biologico preesistente.
Nel primo caso una forma di vita dovrebbe essere in grado di auto-organizzarsi in un sistema in grado di generare informazione.
Sostanzialmente si ritiene che in questo modo la vita ebbe origine sulla Terra, da un brodo primordiale di sostanze chimiche in un ambiente acquoso che ha permesso la generazione di molecole autoreplicanti, che poi sono andate incontro a mutazioni per poi evolvere.
Quindi difronte ad una ipotetica o reale vita alternativa, utile sarebbe porci una serie di domande e cercare risposte che inquadrino la questione in termni di informazione domandandoci:
Le informazioni ereditarie da dove vengono e quante ne sono coinvolte? Un sistema biologico è distinguibile da un sistema chimico perchè possiedono componenti che possono dare origine a molte potenziali combinazioni; a ciò dobbiamo aggiungere che i sistemi biologici possiedono una memoria molecolare (genotipo) che viene modificata nel corso di mutazioni e selezioni, cambiamenti che se permettono all'organismo di sopravvivere possono essere trasferiti alle generazioni successive.
Nel valutare la presenza di potenziali forme di vita alternativa si dovrebbe quindi porre particolare attenzione al materiale genetico, e qui sovviene un altro quesito:
Qual'è il numero minimo di informazioni genetiche necessarie per dare origine a tutti quei meccanimsmi che possano favorire l'ereditabilità di queste informazioni e che possano permettere l'instaurarsi di modificazioni a loro carico che possano aumentare le combinazioni (pool di informazioni genetiche) disponibili?
Ovviamente dipende dal tipo di sistema che andiamo a prendere in considerazione e al tipo di modifiche che subisce nel tempo.
L'obiettivo posto dall'autore è quello di determinare il numero di informazioni ereditabili che possono essere coinvolte, escludendo quelle che non nascono all'interno del sistema.
Ponendo varie domande tra cui:
Riporto l'esempio dell'autore:
Conteggio delle informazioni ereditabili.
Quanti tipi diversi di subunità (x) dovrebbe contenere il materiale genetico? Per un polimero binario x= 2 per uno quaternario come nel caso di un acido nucleico x= A,T /U,G,C.
Quale dovrebbe essere la lunghezza del materiale genetico?
Quali sono le possibili combinazioni che ne potrebbero derivare dalla modificazione di tali informazioni e quali si potrebbero presentare con maggiore probabilità?
Le modificazioni a cui vanno incontro tali componenti genetiche non hanno tutte la stessa probabilità di subire modificazioni e di dare origine alle varie combinazioni che ne possano derivare, ciò è dovuto al fatto che alcune subunità possono non essere presenti nelle giuste quantità o che venga a trovarsi meno frequentemente nel materiale genetico per far si che ciò si verifichi. Alcune reazioni ad esempio possono in ambito chimico e biochimico essere totalmente sfavorevoli, basti citare le lunghe sequenze di G nel DNA. Il numero di possibili combinazioni, seguendo lo schema proposto dall'autore, è x^n.
Se tutte queste combinazioni fossero possibili ognuna avrebbe una probabilita a priori di verificarsi di x^-n e il contenuto informativo, il numero di informazioni associato a particolari combinazioni sarebbe log2 (x^n). Ciò può essere espresso anche come 2#bits = x^n. per un polimero binario, #bits = n; for a nucleic acid polymer, #bits = 2n.
Se le varie combinazioni non hanno la stessa probabilità di verificarsi, allora il contenuto di informazioni associate ad una particolare combinazione realizzata deve essere calcolata sulla base della sua probabilità a priori di verificarsi (PK), che varia da 0 a 1. Il contenuto informativo (numero di bit) associato ad una particolare combinazione è realizzata -log2 (PK).
In linea di principio, quindi, l'autore prende in considerazione due vie attraverso le quali una nuova forma di vita può sorgere: direttamente dalla chimica o dalla scissione di una forma biologica esistente.
Se la vita nasce dalla chimica, per fare un esempio in un modo simile agli eventi che si pensa si siano verificati sulla primitiva Terra,

il sistema inizierebbe da zero informazioni ereditabili, eventuali cambiamenti a seguito di un periodo di chimica prebiotica, attraverso fattori che ne favoriscano una adeguata trasformazione in un eventuale sistema indirizzato ad una probabile creazione della vita, forse raggiungerebbe un elevato livello di complessità chimica, da cui potrebbe derivare una memoria molecolare, la quale potrebbe iniziare un processo di replicazione, mutazione ed eventuale evoluzione.
Se, invece, la vita nasce da un'altra forma di vita, allora può avere un inizio "privilegiato", beneficiando di un ambiente chimico che è stato plasmato da una forma di vita preesistente, basata su un insieme di informazioni preesistenti.
SI pensa che questo sia stato il tipo di transizione che si è verificato sulla Terra quando la vita basata sull'RNA ha dato il via alla vita basata sul DNA e sui complessi proteici.
Questo passaggio probabilmente ha comportato un sostanziale trasferimento di informazioni che erano maturate in molecole di RNA e che sono state riportate sul DNA attraverso processi di trascrizione inversa. Molti biologi "vedono" echi di quelle informazioni ancestrali nelle sequenze di RNA ribosomiale, tRNA, e RNA contemporanei.
È stato suggerito che la vita basata sull'RNA come depositario dell'informazione sia stata preceduta da un'altra forma di vita, forse basata da un molecola depositaria dell'informazione simile ad un acido nucleico, come la molecola TNA (threose nucleic acid) o molecole come GNA (glicole nucleic acid).
Nella biologia moderna comunque non sembrano esserci tracce di una qualche forma di vita pre-RNA.
E'anche possibile che vi fosse vita sulla terra anche prima della formazione di una molecola di RNA e che non vi fosse alcuna somiglianza nelle componenti che permettessero il trasferimento di informazione, con l'RNA. Informazioni genetiche provenienti da una molecola di pre-RNA potrebbero essere state tradotte in RNA attraverso un meccanismo analogo a quello operato dal ribosoma, ma con l'RNA come output anziché imput.
In alternativa, una vita basata su una molecola di pre-RNA può aver facilitato l'emergere della vita basata sull'RNA, ma senza il trasferimento delle informazioni.
Una forma di vita Pre-RNA può aver generato molecole di RNA come un prodotto metabolico, e, infine, le molecole di RNA sono andate incontro a cambiamenti di vario tipo.
La discussione di cui sopra, sottolinea che ci sono quindi molti percorsi possibili attraverso cui una forma di vita può dar luogo ad un altra, ma non affronta la questione di come si possa creare una nuova forma di vita iniziale.
Quando gli astronomi parlano di zone abitabili si riferiscono ad un'orbita planetaria che è ad una distanza adeguata da una stella per mantenere acqua liquida sulla superficie dei pianeti. Questa potrebbe essere una definizione troppo restrittiva o troppo generosa, a seconda dei punti di vista.
Forse la vita può esistere in un ambiente non acquoso, anche se ci sono pochi dati a sostegno di questa congettura.
Al contrario, anche un laghetto temperato povero di sostanze organiche diluite può essere insufficiente per permettere la formazione della vita. Lo stagno avrebbe bisogno di accumulare eteropolimeri di composizione variabile, alcuni dei quali dovrebero essere in grado di raggiungere una complessita tale da iniziare processi di replicazione e fornire la base per la memoria molecolare. La possibilità che ciò si verifichi non è ancora quantificabile e non è chiara se si tratti di un evento comune o estremamente raro.
L'autore inoltre espone un recente studio portato avanti dal suo team cercando, riproducendolo a tavolino, di descrivere un esempio di sistema chimico, che può subire evoluzione darwiniana in un modo autosufficiente . "Auto-sostenuta" in questo contesto significa che tutte le informazioni necessarie al sistema per essere sottoposto ai processi di evoluzione sono parte del sistema che si evolve, non provengono dall'esterno da un altra forma di vita per intenderci. Il sistema chimico coinvolge coppie di ribozimi che catalizzano reciproca sintesi unendo insieme due blocchi oligonucleotidici (Figura a lato sotto).
Nell'immagine sottostante in A è possibile osservare lo schema che mostra un ciclo di replicazione accoppiato di ribozimi (E in blu ed E' in arancione) che catalizzano una reciproca sintesi unendo due substrati oligonucleotidici corrispondenti, con A' e B' per formare E' e A con B per formare E. Ogni substrato contiene sei nucleotidi di sequenza variabile che sono riconosciuti dall'enzima. La struttura terziaria è basata sul modello omologo alla struttura cristallina dell'enzima RNA ligasi I.
IN B è mostrata la sequenza e la struttura secondaria del complesso E-A'-B' con le regioni genetiche racchiuse da un rettangolo e le regioni corrispondenti al dominio funzionale dell'enzima indicati in colore e con i nucleotidi immutabili che sono fondamentali per la replicazione (sono mostrati in nero). Inoltre nell'immagine in B potete notare una freccia ricurva, sta ad indicare il sito di legame di A' e B' per formare E.
Ciascuno dei due blocchi può adottare migliaia di composizioni alternative possibili e quindi formare in teoria milioni di combinazioni diverse. Ognuna delle tante combinazioni possibili potrebbe essere in grado di auto-replicarsi e trasmettere informazioni sulla composizione della sua progenie. Varianti che si replicano in maniera più efficiente possono crescere fino ad arrivare a dominare la popolazione, fino all'adozione di nuove varianti più vantaggiose che soppiantano i loro predecessori in una battaglia senza fine per la sopravvivenza
La popolazione di ribozimi costituisce un sistema di sintesi genetica, ma non è una nuova forma di vita, infatti come fa notare l'autore presenta due grandi limitazioni.
Primo, le molecole contengono solo 24 bit (12 coppie di basi) di informazioni ereditabili per codificare una specifica funzione. Secondo, la replica dipende da 60 bit (30 nucleotidi definiti) che però sono stati forniti all'inizio e non sono soggetti a mutazione e selezione (Figura 3B). Così degli 84 bit totali necessari al sistema per iniziare a replicare ed evolvere, solo un quarto può essere considerato come parte della memoria molecolare del sistema che può essere soggetto a cambiamenti. In questo caso il sistema di sintesi genetica non è una nuova forma di vita perché opera per lo più su informazioni (bit) presi in prestito. Insomma non contiene abbastanza bit (informazioni) ereditabili rispetto a quelle per avviare il funzionamento di tutto il sistema per proseguire il suo percorso.
Conclude Joyce"Forse la prima vera forma di vita alternativa alla biologia terrestre si troverà su un pianeta extrasolare, in una roccia proveniente da Marte, o all'interno di un ambiente estremo sulla Terra.
Più probabilmente, sarà l'opera di una specie intelligente che ha scoperto i principi dell'evoluzione darwiniana e ha imparato a concepire sistemi chimici che hanno la capacità di generare informazioni genetiche per conto proprio come l'uomo. "
Fonti:
http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.1001323
Nota:
Ero davvero indeciso se parlare di questo saggio pubblicato da Joyce su Plos biology, mi rendo conto che forse l'argomento trattato possa risultare pesante e non di immediata comprensione, che spero non aver reso ancora più difficile a causa del mio modo di esporverla; vi assicuro che mi sono impegnato al massimo per rendervela semplice...ma non troppo :)... ovviamente sperando di non aver travisato nulla nella traduzione e nella comprensione del testo...
In tal caso dateci sotto! Alla prossima.



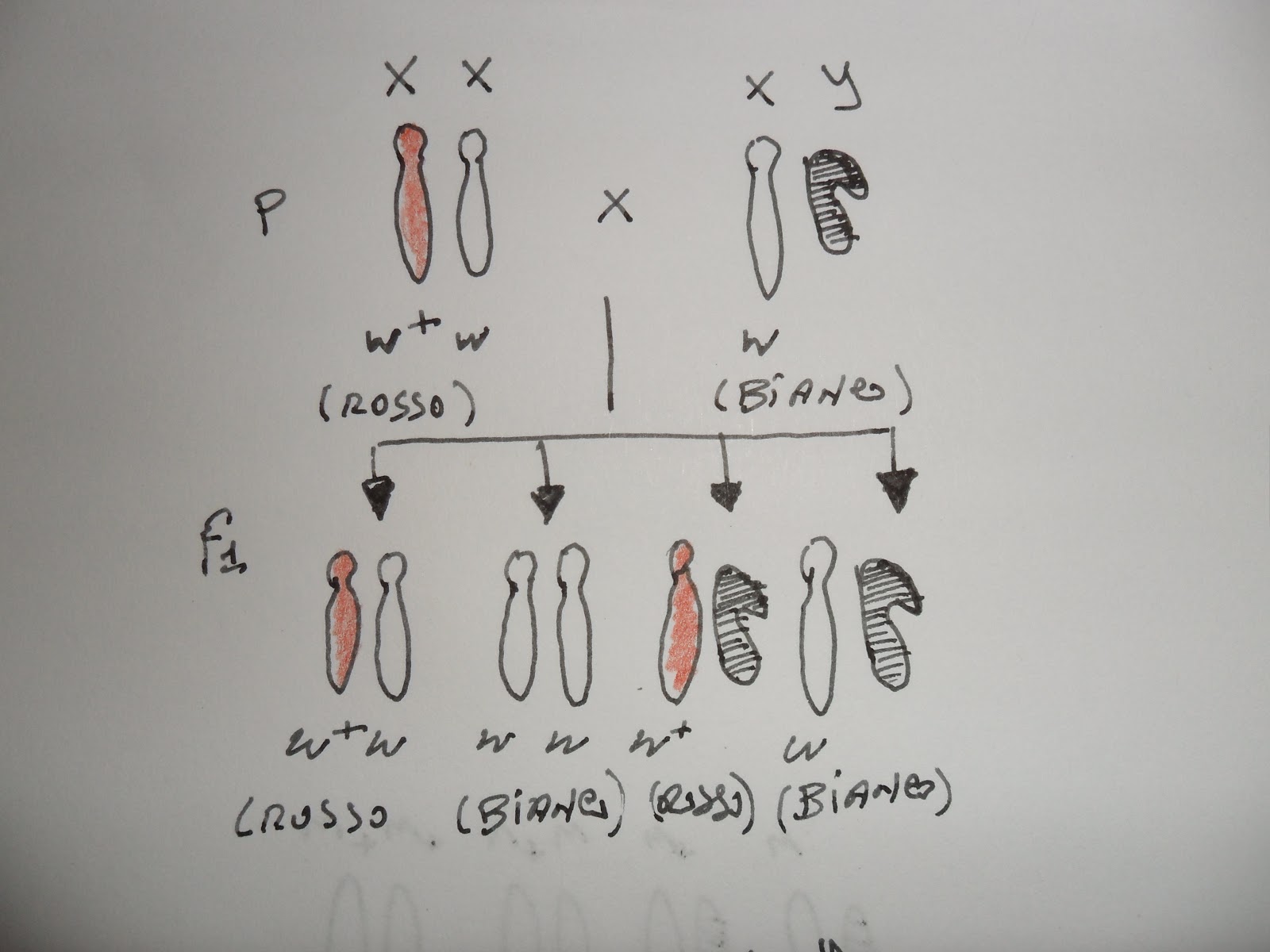
 Infatti riuscì a dimostrare citologicamente (vedere immagine a lato e sotto) che le femmine con occhio bianco ottenute dall'incrocio con maschio occhi rossi, avevano due cromosomi X e un cromosoma Y, mentre metà dei maschi con occhio bianco mostrava un unico cromosoma X e due Y.
Infatti riuscì a dimostrare citologicamente (vedere immagine a lato e sotto) che le femmine con occhio bianco ottenute dall'incrocio con maschio occhi rossi, avevano due cromosomi X e un cromosoma Y, mentre metà dei maschi con occhio bianco mostrava un unico cromosoma X e due Y.