Verso la fine degli anni 40 due scienziati, Joshua Lederberg e Edward Tatum, analizzarono due ceppi di E.coli, entrambi auxotrofi complessi e fecero la sorprendente scoperta che i geni si trasferiscono da una cellua all'altra di E.coli.
Nessuno dei due ceppi era in grado di crescere su un terreno minimo, il ceppo A necessitava di metionina e biotina, il ceppo B di un supplemento di treonina, leucina, tiamina...
A breve il post (chiedo scusa ma blogger ha fatto i capricci tutto il giorno...)
giovedì 29 novembre 2012
mercoledì 28 novembre 2012
LA GENETICA DEI BATTERI: La trasformazione.
Avete mai sentito parlare di trasferimento genico orizzontale? Questa espressione significa che i caratteri coinvolti non sono trasferiti in modo ereditario da una generazione all'altra , ma provengono da individui non correlati o a specie differenti.
I batteri possono trasferire geni da un ceppo ad un altro attraverso tre meccanismi differenti, noti rispettivamente come trasformazione, coniugazione e trasduzione.
Questi meccanismi di trasferimento genico sono accomunati dalla presenza di un donatore, l'organismo che fornisce il materiale genetico che sarà trasferito e da un ricevente il quale ovviamente riceverà il materiale genetico dal donatore. I batteri che fungono da riceventi, a seconda del meccanismo di trasferimento genico che subiscono sono chiamati rispettivamente: trasformanti, ex-coniuganti, trasduttanti.
La trasformazione.
Questo processo è unidirezionale, va solo dal donatore al ricevente, inoltre la maggior parte dei riceventi riceve solo una piccola percentuale del DNA del donatore.
Vi sono molte specie batteriche che vanno incontro ad una trasformazione naturale, tra cui lo Streptococcus pneuomoniae, Neisseria gonorrhoeae agente eziologico della gonorrea ecc...
Questo fenomeno rappresenta la captazione da parte di una cellula di una molecola di DNA nudo dal mezzo circostante e la sua incorporazione all'interno del ricevente.
Il processo è del tutto casuale e la porzione qualsiasi di un genoma può essere trasferita tra procarioti.
Quando i batteri vanno incontro a lisi lasciano una grande quantità di materiale genomico nell'ambiente esterno, frammenti di dimensioni diverse che possono a seconda dei casi contenere svariati geni, se uno di questi frammenti si pone in contatto con una cellula definita competente (la cellula ricevente) cioè una cellula in grado di poter integrare il materiale genomico, tale frammento potrà essere trasportato all'interno della cellula e integrato nel suo genoma. Non tutte le cellule procariote vanno incontro a questo processo, non tutte sono competenti; ciò che rende una cellula competente sono un insieme di fattori particolarmente complessi e per far si che una cellula competente possa assumere DNA attraverso questo processo vi è bisogno che si verifichino specifiche condizioni, ad esempio il batterio che funge da ricevente deve trovarsi in un determinato stadio di crescita.
Quando tali microrganismi hanno raggiunto lo stadio di crescita adeguato producono specifiche proteine che vengono definite fattori di competenza fondamentali per la produzione di altre molecole proteiche richieste per la trasformazione. La trasformazione che avviene per via naturale è stata osservata solo in poche specie gram-negative e gram positive.
Il meccanismo della trasformazione è stato ampiamente studiato nello S.pneumoniae, Un frammento di DNA a doppio filamento grande abbastanza si lega ad una cellule competente, il processo è casuale e i frammenti donatori competono fra di loro, poi il DNA è scisso dalle endonucleasi in porzioni a doppio filamento di dimensioni comprese tra 5 e 15 chilobasi. La captazione richiede una spesa energetica non indifferente, un frammento viene idrolizzato da esonucleasi, associata all'involucro nel corso della captazione, mentre l'altro si unisce a piccole proteine e si sposta attraverso la membrana plasmatica. Il frammento a singolo filamento può quindi allinearsi ad una regione omologa del genoma e venire integrato.
In una coltura di laboratorio il numero di batteri competenti dipenderà dalle condizioni di crescita, in genere le cellule che mostrano la maggiore probabilità di essere competenti sono quelle in attiva divisione cellulare. Queste cellule si vengono a trovare nel periodo di crescita esponenziale avvicinandosi alla regione di plateu, momento nel quale i nutrienti fondamentali ai microrganismi iniziano ad essere limitanti per la crescita.
La trasformazione è un meccanismo utilizzato ampiamente per la mappatura genica. Le molecole di DNA estratte dallo Streptococcus si rompono in circa 50 frammenti, quindi ciascun frammento di DNA rappresenta circa il 2% del genoma.
I batteri possono trasferire geni da un ceppo ad un altro attraverso tre meccanismi differenti, noti rispettivamente come trasformazione, coniugazione e trasduzione.
Questi meccanismi di trasferimento genico sono accomunati dalla presenza di un donatore, l'organismo che fornisce il materiale genetico che sarà trasferito e da un ricevente il quale ovviamente riceverà il materiale genetico dal donatore. I batteri che fungono da riceventi, a seconda del meccanismo di trasferimento genico che subiscono sono chiamati rispettivamente: trasformanti, ex-coniuganti, trasduttanti.
La trasformazione.
Questo processo è unidirezionale, va solo dal donatore al ricevente, inoltre la maggior parte dei riceventi riceve solo una piccola percentuale del DNA del donatore.
Vi sono molte specie batteriche che vanno incontro ad una trasformazione naturale, tra cui lo Streptococcus pneuomoniae, Neisseria gonorrhoeae agente eziologico della gonorrea ecc...
Questo fenomeno rappresenta la captazione da parte di una cellula di una molecola di DNA nudo dal mezzo circostante e la sua incorporazione all'interno del ricevente.
Il processo è del tutto casuale e la porzione qualsiasi di un genoma può essere trasferita tra procarioti.
Quando i batteri vanno incontro a lisi lasciano una grande quantità di materiale genomico nell'ambiente esterno, frammenti di dimensioni diverse che possono a seconda dei casi contenere svariati geni, se uno di questi frammenti si pone in contatto con una cellula definita competente (la cellula ricevente) cioè una cellula in grado di poter integrare il materiale genomico, tale frammento potrà essere trasportato all'interno della cellula e integrato nel suo genoma. Non tutte le cellule procariote vanno incontro a questo processo, non tutte sono competenti; ciò che rende una cellula competente sono un insieme di fattori particolarmente complessi e per far si che una cellula competente possa assumere DNA attraverso questo processo vi è bisogno che si verifichino specifiche condizioni, ad esempio il batterio che funge da ricevente deve trovarsi in un determinato stadio di crescita.
Quando tali microrganismi hanno raggiunto lo stadio di crescita adeguato producono specifiche proteine che vengono definite fattori di competenza fondamentali per la produzione di altre molecole proteiche richieste per la trasformazione. La trasformazione che avviene per via naturale è stata osservata solo in poche specie gram-negative e gram positive.
Il meccanismo della trasformazione è stato ampiamente studiato nello S.pneumoniae, Un frammento di DNA a doppio filamento grande abbastanza si lega ad una cellule competente, il processo è casuale e i frammenti donatori competono fra di loro, poi il DNA è scisso dalle endonucleasi in porzioni a doppio filamento di dimensioni comprese tra 5 e 15 chilobasi. La captazione richiede una spesa energetica non indifferente, un frammento viene idrolizzato da esonucleasi, associata all'involucro nel corso della captazione, mentre l'altro si unisce a piccole proteine e si sposta attraverso la membrana plasmatica. Il frammento a singolo filamento può quindi allinearsi ad una regione omologa del genoma e venire integrato.
In una coltura di laboratorio il numero di batteri competenti dipenderà dalle condizioni di crescita, in genere le cellule che mostrano la maggiore probabilità di essere competenti sono quelle in attiva divisione cellulare. Queste cellule si vengono a trovare nel periodo di crescita esponenziale avvicinandosi alla regione di plateu, momento nel quale i nutrienti fondamentali ai microrganismi iniziano ad essere limitanti per la crescita.
La trasformazione è un meccanismo utilizzato ampiamente per la mappatura genica. Le molecole di DNA estratte dallo Streptococcus si rompono in circa 50 frammenti, quindi ciascun frammento di DNA rappresenta circa il 2% del genoma.
Negli esperimenti di mappatura genica che utilizza la trasformazione , due geni vengono definiti associati se si trovano sullo stesso frammento di DNA.
In questo caso dobbiamo sottolineare che non si deve parlare di associazione allo stesso modo di quando ci riferiamo agli eucarioti, in cui tutti i geni che manifestano una percentuale inferiore al 50% di ricombinazione sono definiti come associati.
Se due geni di nostro interesse saranno associati tra di loro si troveranno molto frequentemente sullo stesso frammento di DNA e le cellule avranno una elevata possibilità di essere co-trasformate da essi, se invece tali geni sono distanti, tanto più sarà bassa la possibilità che essi si trovino sullo stesso frammento genico.
Esempio:
Si vuole determinare se due geni sono tra di loro associati, vengono usate tre diverse classi di molecole donatrici di DNA: a+b-; a-b+; a+b+. Le cellule riceventi sono caratterizzate dall'essere tutte a-b-.
In diversi esperimenti sono utilizzate quantità decrescente di molecole di DNA donatrici a+b- e a-b+ per dimostrare la frequenza di trasformazione singola di a-b- ; a+b- e a-b+. In teoria la frequenza di trasformanti dovrebbe diminuire al decrescere della concentrazione di DNA del donatore.
La frequenza della doppia trasformazione a-b- verso a+b+ utilizzando il DNA del donatore a+b+ viene poi determinata come funzione della concentrazione decrescente di DNA.
Se i due geni sono strettamente associati tra di loro, quindi se si vengono a trovare con una frequenza elevata sullo stesso frammento del DNA sarà attesa una curva con lo stesso andamento della singola trasformazione, in quanto la doppia trasformazione come quella singola richiede l'incorporazione di un singolo frammento di DNA.
Se i due geni sono molto distanti tra di loro, e quindi non avranno una elevata probabilità di essere presenti sullo stesso frammento di DNA, la doppia trasformazione può avvenire solo se due frammenti distinti di DNA entrano all'interno della cellula che funge da ricevente e avvengono due eventi distinti di ricombinazione.
Quindi se la concentrazione del DNA diminuisce di dieci volte, la frequenza di doppia trasformazione deve diminuire di cento volte.
Ad esempio se la concentrazione di DNA diminuisce da 1,0 a 0,1 unità per ml, la frequenza di singola trasformazione diminuisce di dieci volte, da 1,0 a 0,1 cellule per unità di volume; la frequenza di doppia trasformazione per due geni su due frammenti differenti scende a 0,1x0,1=0,01 cellule per unità di volume, un calo di cento volte.
Quindi analizzando le frequenze di singola e doppia trasformazione esclusivamente come funzioni della diminuzione della concentrazione di DNA si può individuare con sicurezza l'associazione.
Altro esempio: in un batterio trasformato, vogliamo stabilire l'ordine dei geni, si estrae il DNA da batteri donatori, il DNA si frammenta, su ognuno di questi frammenti ci saranno i geni di nostro interesse, come può essere notato nell'immagine nei genotipi dei trasformanti non troviamo mai il gene a ed il gene c, segno che sono i più distanti quindi il gene b è al centro.

Se due geni di nostro interesse saranno associati tra di loro si troveranno molto frequentemente sullo stesso frammento di DNA e le cellule avranno una elevata possibilità di essere co-trasformate da essi, se invece tali geni sono distanti, tanto più sarà bassa la possibilità che essi si trovino sullo stesso frammento genico.
Esempio:
Si vuole determinare se due geni sono tra di loro associati, vengono usate tre diverse classi di molecole donatrici di DNA: a+b-; a-b+; a+b+. Le cellule riceventi sono caratterizzate dall'essere tutte a-b-.
In diversi esperimenti sono utilizzate quantità decrescente di molecole di DNA donatrici a+b- e a-b+ per dimostrare la frequenza di trasformazione singola di a-b- ; a+b- e a-b+. In teoria la frequenza di trasformanti dovrebbe diminuire al decrescere della concentrazione di DNA del donatore.
La frequenza della doppia trasformazione a-b- verso a+b+ utilizzando il DNA del donatore a+b+ viene poi determinata come funzione della concentrazione decrescente di DNA.
Se i due geni sono strettamente associati tra di loro, quindi se si vengono a trovare con una frequenza elevata sullo stesso frammento del DNA sarà attesa una curva con lo stesso andamento della singola trasformazione, in quanto la doppia trasformazione come quella singola richiede l'incorporazione di un singolo frammento di DNA.
Se i due geni sono molto distanti tra di loro, e quindi non avranno una elevata probabilità di essere presenti sullo stesso frammento di DNA, la doppia trasformazione può avvenire solo se due frammenti distinti di DNA entrano all'interno della cellula che funge da ricevente e avvengono due eventi distinti di ricombinazione.
Quindi se la concentrazione del DNA diminuisce di dieci volte, la frequenza di doppia trasformazione deve diminuire di cento volte.
Ad esempio se la concentrazione di DNA diminuisce da 1,0 a 0,1 unità per ml, la frequenza di singola trasformazione diminuisce di dieci volte, da 1,0 a 0,1 cellule per unità di volume; la frequenza di doppia trasformazione per due geni su due frammenti differenti scende a 0,1x0,1=0,01 cellule per unità di volume, un calo di cento volte.
Quindi analizzando le frequenze di singola e doppia trasformazione esclusivamente come funzioni della diminuzione della concentrazione di DNA si può individuare con sicurezza l'associazione.
Altro esempio: in un batterio trasformato, vogliamo stabilire l'ordine dei geni, si estrae il DNA da batteri donatori, il DNA si frammenta, su ognuno di questi frammenti ci saranno i geni di nostro interesse, come può essere notato nell'immagine nei genotipi dei trasformanti non troviamo mai il gene a ed il gene c, segno che sono i più distanti quindi il gene b è al centro.

lunedì 26 novembre 2012

giovedì 22 novembre 2012
MAPPA GENETICA E POSIZIONE FISICA DEI GENI.
La distanza fisica reale tra due geni non mostra sempre una correlazione con la distanza della mappa genetica.
Il fatto che non vi sia una elevata correlazione tra la frequenza di ricombinazione e la distanza fisica tra i geni su un cromosoma è dovuto a molti fattori tra i quali l'esistenza di doppi, tripli o multipli crossing over.
Ne consegue quinidi che la possibilità che avvenga un doppio crossing over tra due geni molto distanti tra di loro su un cromosoma è elevata.
Inoltre come abbiamo accennato nei post precedenti, la frequenza massima di ricombinazione di un incrocio è del 50% o inferiore.
Un fattore che può ridurre l'accuratezza della frequenza di ricombinazione come misura atta a rilevare le distanze cromosomiche, è proprio il fatto che due geni che sono distanti tra di loro su un cromosoma non ricombineranno mai più del 50%; inoltre la ricombinazione è un processo che non avviene uniformemente per tutta la lunghezza del cromosoma, esistono i cosidetto "hot spots" i punti caldi; possiamo definirli come dei siti lungo un cromosoma dove il processo di ricombinazione si verifica più frequentemente, come se fosse facilitato in quel punto la ricombinazione, e regioni di un cromosoma dove la ricombinazione avviene molto più raramente come nelle vicinanze del centromero (deserti).
Da quando sono state iniziate le mappature i genetisti hanno generato delle equazioni matematiche definite come funzioni di mappa.
Sono state ideate per compensare queste discrepanze che abbiamo solo vagamente accenato sopra tra le vere distanze fisiche e le frequenze di ricombinazione.
Queste funzioni introducono sensibili cambiamenti nelle frequenze di ricombinazione tra geni molto distanti tra di loro e cambiamenti minimi elle distanze di mappa di geni molto vicini.
E'stato ossevato però che le correzioni per distanze maggiori sono comunque imprecise, in quanto le funzioni di mappa si basano su presupposti come assenza di interferenza (ne parliamo in un altro post) e raramente ciò non avviene nella realtà.
In sintesi le funzioni di mappa hanno lo scopo di ridurre le discrepanze tra distanza fisica e frequenza di ricombinazione, ma il metodo migliore per produrre una mappa accurata è ancora quello di sommare intervalli piccoli, mappando i geni che sono molto distanti tra di loro erificando che essi sono associati con lo stesso gruppo di geni presenti tra i due geni distanti.
A suo volta questo cosa comporta? Le mappe genetiche più accurate sono quelle che si ottengono mediante un grande numero di incroci le quali però devono essere continuamente rifinite, su un cromosoma infatti non sono presenti solo due o tre geni, ma molti di più, quindi man mano che si scoprono le mappe devono essere rifinite.
Le frequenze di ricombinazione non sono neanche identiche per tutte le specie, ne consegue quindi che per un organismo 1 u.m. può corrispondere a migliaia di coppie di basi, mentre per un altro organismo a centomila.
mercoledì 21 novembre 2012
LA RICOMBINAZIONE: il crossing over separa i geni associati.
Cosa avviene durante la ricombinazione?

Come possiamo vedere nell'immagine il crossing over determina uno scambio di materiale genetico, unisce i quattro cromatidi omologhi ed è fondamentale per l'adeguata segregazione dei cromosomi nelle successive divisioni meiotiche.
Nell'immagine, partendo dall'alto, abbiamo una coppia di cromosomi omologhi che sono stati duplicati allinizio della profase della meiosi I.
Durante il leptotene e lo zigotene della provasi I il complesso sinaptinemale facilita l'allineamento delle regioni corrispondenti dei cromosomi omologhi permettendo la ricombinazione.
I chiasmi sono i siti dove il crossing over è visibile.
Quando il complesso sinaptinemale si disassembla durante il diplotene i cromosomi omologhi restano appaiati a livello dei chiasmi.
Quando i due omologhi si separano in anafase I iniziando dai centormeri, le estremità dei due cromatidi ricobinanti si allontanano dai rispettivi cromatidi fratelli e i chiasmi, dalla loro posizione d'origine si muovono verso l'estremità del cromosoma o telomero. Questo processo viene definito di terminalizzazione. La meiosi procede alla fine producendo quattro cellule aploidi contenente un cromosoma a testa, grazie a questo processo i cromosomi omologhi si sono scambiati dei pezzi.
Morgan era convinto che i chiasmi rappresentassero i siti di crossing over tra i cromosomi, e che tale fenomeno determinasse la ricombinazione.
Ciò lo convinse a sostenere una ipotesi, coppie diverse di geni mostravano livelli di associazione differenti in quanto i geni sono disposti in sequenza lineare lungo i cromosomi, più due geni si troveranno vicini su un cromosoma, minore possibilità si avrà che essi vengano separati da un processo che tagli e ricombini la sequenza lineare dei geni. In parole povere se si assume la possibilità che i chiasmi i possano formare su qualsiasi porzione del cromosoma con la stessa probabilità, allora si la probabilità che avvenga un crossing over tra due geni auenta con l'aumentare dela distanza tra di essi.
Sturtevant ebbe una idea, considerare la percentuale di ricombinazione, definita anche come sequenza di ricombinazione, come indice della distanza fisica che separa due geni qualsiasi sullo stesso cromosoma. Presa come unità percentuale in maniera arbitraria lingo il cromosoma, essa venne chiamata centimorgan (cM) definita anche come unità di mappa (u.m).
Esempio, se da un incrocio si osserva che i geni per il colore dell'occhio (w) e per il colore del corpo (g) ricombinano nell'1,1% dei casi essi sono distanti 1,1 u.m.
Come unità di misura, l'unità di mappa è un indice di probabilità di ricombinazione che ci aiuta a determinare la distanza tra i geni. I genetisti hanno sfruttato questo concetto per mappare migliaia di geni di Drosophila.
Se l'associazione tra geni è definita dal fatto di avere una proporzione di classi ricombinanti più bassa di quella delle classi parentali, allora una frequenza di ricombinazione inferiore al 50% indica associazione.
I ricombinanti possono essere presenti in numero maggiori degli individui che mostrano genotipo parentale? Come determiniamo la posizione dei geni quando i parentali sono presenti in numero quasi uguale ai ricombinanti?
Quando da un incrocio abbiamo una progenie che presenta il 50% dei ricombinanti sarebbe più corretto parlare di riassortimento.
Quando due geni sono localizzati su due cromosomi differenti , quindi non omologhi, seguiranno la legge di mendel dell'assortimento indipendente, in quanto i due cromosomi possono allinearsi sul fuso meiotico primo in uno dei qualsiasi di due orientamenti equiprobabili. In questo modo un diibrido fra questi due geni produrrà quattro possibili gameti, AB, Ab,aB,ab con una frequenza approssimativamente identica.
Nonostante questo geni presenti sugli stessi cromosomi ma molto distanti gli uni dagli altri possono presentare una ricombinazione che sfiora il 50%.
Non sono mai stati osservate frequenze di ricombinazione tra due geni maggiori del 50%, quindi i ricombinanti discendenti da un incrocio ad esempio una generazione F2 non sono mai la maggioranza.
Non sono mai stati osservate frequenze di ricombinazione tra due geni maggiori del 50%, quindi i ricombinanti discendenti da un incrocio ad esempio una generazione F2 non sono mai la maggioranza.
Questo limite dipende fondamentalmente da due aspetti dei cromosomi durante la meiosi I.
Primo fra due geni lontani possono avvenire eventi multipli di crossing over, secondo la ricombinazione avviene quando ciascun cromosoma è giò replicato in due cromatidi.
Quindi le frequenze di ricombinazione del 50% suggeriscono che i geni si possono trovare o su cromosomi differenti oppure che sono sullo stesso cromosoma, ma che possono trovarsi molto distanti tra di loro e quindi essere coinvolti in processi multipli di crossing over.
Come si capisce se due geni sono sullo stesso cromosoma?
Bisogna programmare degli incroci che ci permettano di verificare l'associazione tra due geni , situati sullo stesso cromosoma e tra di essi.
Anche se gli incroci tra due geni che si trovano distanti gli uni dagli altri, potrebbero indicare che non ci sta associazione, in quanto i parentali e i ricombinanti, si può dimostrare che essi sono presenti sullo stesso cromosoma solo se si individuano uno o più geni intermedi che mostrano associazione con entrambi.
Il crossing over è il meccanismo che permette il verificarsi della ricombinazione, tramite questo processo i geni presenti sui cromosomi materni si spostano su quello paterno e viceversa, quando gli omologhi si scambiano parti durante la prima divisione meiotica. I chiasmi sono la manifestazione visibile di questi scambi, e dal momento che la formazione dei chiasmi può avvenire ovunque sul cromosoma più due geni sono distanti su un cromosoma maggiore probabilità che tra di essi si formino chiasmi.
Ecco perchè a frequenze di ricombinazione elevate corrispondono a distanze elevate tra i geni.
Per quanto detto prima le frequenze di ricombinazione variano tra lo 0% e il 50%; valori minori del 50% indicano che due geni sono associati, e quindi localizzati sullo stesso cromosoma, geni che invece mostrano una frequenza del 50% indicano che due geni sono presenti su cromosomi diversi o perchè sono sullo stesso cromosoma ma molto distanti l'uno dall'altro.
lunedì 19 novembre 2012
LA RICOMBINAZIONE.
La ricombinazione, processo che avviene durante la meiosi, quando il crossing over separa i geni associati...
tra poche ore il post.
tra poche ore il post.
sabato 17 novembre 2012
DIBATTITO SCIENZA.

L'iniziativa deriva da un gruppo di blogger, giornalisti scientifici, vedasi Marco Ferrari giornalista e blogger di Leucophaea, o il blogger Moreno Colaiacovo di mygenomix ,e semplici appassionati di scienza, i quali si sono dati appuntamento su Facebook, dal dibattito iniziato è scaturita l'idea di porre sei domande ai politici, ai candidati del centrosinistra per ora ma non è da escludere che verranno posti anche ai rappresentanti di altre fazioni politiche, riguardanti temi come gli Ogm, legge 40 ecc...
L'iniziativa e tutto ciò che ne è da cornice potete visualizzarla al seguente indirizzo Facebook "Dibattito scienza".
Anche il giornale Le Scienze ne parla nella home page (dibattito scienza).
Le domande sono le seguenti:
1. Quali politiche intende perseguire per il rilancio della ricerca in Italia, sia di base sia applicata, e quali provvedimenti concreti intende promuovere a favore dei ricercatori più giovani?
2. Quali misure adotterà per la messa in sicurezza del territorio nazionale dal punto di vista sismico e idrogeologico?
3. Qual è la sua posizione sul cambiamento climatico e quali politiche energetiche si propone di mettere in campo?
4. Quali politiche intende adottare in materia di fecondazione assistita e testamento biologico? In particolare, qual è la sua posizione sulla legge 40?
5. Quali politiche intende adottare per la sperimentazione pubblica in pieno campo di OGM e per l’etichettatura anche di latte, carni e formaggi derivati da animali nutriti con mangimi OGM?
6. Qual è la sua posizione in merito alle medicine alternative, in particolare per quel che riguarda il rimborso di queste terapie da parte del SSN?
Alle domande per ora sembra aver risposto solo Laura Puppato.
Al seguente link la risposta da lei data alle domande, inoltre fate anche una capatina su Leucophaea per maggiori informazioni.
Diffondete e partecipate in prima persona a questa iniziativa!!!
CARNEVALE DELLA BIODIVERSITA': IL RITORNO!!!
La notizia l'ho postata su Biosproject: earth, dove troverete parte del bando e il link per andare alla pagina di uno dei tre creatori del carnevale dove potrete trovare tutte le istruzioni per parteciparvi!!!
CARNEVALE DELLA BIODIVERSITA'
CARNEVALE DELLA BIODIVERSITA'
giovedì 15 novembre 2012
ASSOCIAZIONE GENETICA. parte 2
Nel post precedente (associazione genetica) abbiamo notato che vi era nella generazione F2 una preponderanza di individui con genotipo parentale, con notevole spostamento dal rapporto 1:1:1:1 che dovremmo attenderci (per maggiore chiarimento vedere il post ASSOCIAZIONE GENICA: parte 1)



Effettuando una seconda serie di incroci possiamo avere risposta alla nostra domanda.
Incrociamo femmine con occhio rosso e corpo grigio (w+g+/w+g+) con maschi che presentano occhi bianchi e corpo viola w g/Y
Rispettivamente daranno origine a gameti w+g+ e w g.
Le femmine F1 sono diibridi w+g+/wg come nel primo esperimento di incrocio, csa si può dire dei maschi della F1? in che rapporto sono i quattro possibili gameti delle femmine della F1?
Come mostrato nell'immagine sotto, w+ g e w g+ sono i ricombinanti che rappresentano l'1% del totale
mentre w g e w+g+ sono le combinazioni parentali che rappresentano il 99% del totale.
Qui ci sta qualcosa da osservare soprattutto se prendiamo in considerazione gli incroci mostrati nel precedente post e questo.
Come mostrato le frequenze osservate nei vari tipi di progenie dipendo dalla disposizine originale degli alleli nelle femmine F1.
Le combinazioni che si presentano con frequenza particolarmente elevata nella generazione F2 sono le classi patentali e cioè quelle presenti nella generazione P.
Le classi ricombinanti sono presenti con frequenza minore. Le classi parentali e ricombinanti in questo esempio sono opposte a quelle mostrate nel post precedente, in quanto le femmine F2 hanno ricevuto dai genitori combinazioni alleliche diverse dei geni per il colore degli occhi e del corpo. Ne consegue che le percentuali dei tipi ricombinanti e parentali nella progenie sono esatamente le stesse in entrambi gli esperimenti, dimostrando che la frequenza di ricombinazione non dipende dal particolari combinazioni di alleli.
Quando i geni assortiscono in maniera indipendente, il numero degli individui parentali e ricombinanti nella progenie F2 è identico, in quanto ogni individuo F1 doppio eterozigote produce i quattro gameti con la stessa frequenza.
Due geni si dicono associati quando nella progenie F2 il numero di individui con genotipo parentale è molto più elevato di quelli ricombinanti.
Questi geni invece di assortire indipendentemente si comportano come se fossero legati gli uni agli altri, i geni del colore degli occhi e del corpo sono un esempio, infatti le percentuali mostrano che le combinazioni parentali degli alleli w+g e wg+ dell'esempio visto nel precedente post e w+g+ e wg sono mescolate per dare origine a ricombinanti solo in un gamete su 100.
Non sempre però l'associazione è così stretta. Infatti dobbiamo tenere presente che a seconda dei geni che andiamo a prendere in considerazione la percentuale delle classi ricombinanti e parentali cambierà.
Esempio: incrociamo femmine con occhi rossi e ali normali (w+a+/w+a+) con maschi aventi occhi bianchi e ali piccole (w a/Y).

La generazione F1 presenta moscerini con occhi rossi e ali normali; w+a+/w a per le femmine e w+a+/Y per i maschi.
Incrociandoli otteniamo una F2, i genotipi sono mostrati sotto, dove il 65% è di tipo parentale e il 35% di tipo ricombinante.
La percentuale di individi con genotipo parentale è sempre elevata segno che i geni per questi caratteri sono associati, ovviamente paragonando tale percentuale a quella del 99% degli esempi precedenti, l'associazione tra i geni w e m non è così elevata. Le combinazioni parentali per il colore dell'occhio e la grandezza delle ai sono rimescolate in circa 35 gameti su 100 di gran lunga superiore come percentuale a quella dell'1% degli esempi precedenti.
ASSOCIAZIONE GENETICA: una piccola annotazione sul post...
Un lettore mi ha posto una domanda riguardante il post sull'associazione genetica: ciò che dico vale per tutti gli altri lettori che sono passati per questo post e si sono posti lo stesso problema.
Qualora notiate dei problemi, o qualora aveste dei dubbi riguardo ciò che ho scritto non esitate a farmelo sapere...
Scusa il genotipo del maschio "occhi rossi,corpo viola" perchè è w+g/Y? non dovrebbe essere w+w+gg? La "y" cosa indica,hai precisato semplicemente che è maschio???
Qualora notiate dei problemi, o qualora aveste dei dubbi riguardo ciò che ho scritto non esitate a farmelo sapere...
Scusa il genotipo del maschio "occhi rossi,corpo viola" perchè è w+g/Y? non dovrebbe essere w+w+gg? La "y" cosa indica,hai precisato semplicemente che è maschio???
Fate attenzione alle lettere minuscole e maiuscole. Le lettere minuscole si riferiscono al gene, mentre quelle maiuscole al gene, in questo caso la lettera maiuscola Y sta ad indicare il cromosoma Y che non porta geni ne per il colore degli occhi ne per il colore del corpo. Prestate attenzione anche al simbolo della sbarra (/)che viene utilizzato per separare i geni sui cromosomi omologhi, nel nostro esempio X e Y, ma è un ragionamento che può valere per qualsiasi altro cromosoma omologo, ad esempio una coppia di cromosomi X o di qualsiasi altro autosoma; per cui w+g/Y rappresenta il genotipo del maschio che ha un solo cromosoma X che porta gli alleli w e g e un cromosoma Y su cui tali alleli non sono presenti.
Per il suddetto ragionamento avrei potuto mettere per la femmina w g+/w g+ ad esempio, in quanto questi geni sono presenti su entrambi i cromosomi X.
La risposta quindi è si la sbarra l'avevo inserita solo per precisare che è maschio per sottolineare la presenza di questi geni solo sul cromosoma X e non sul cromosoma Y; mi rendo conto che potrebbe creare confusione ad una prima occhiata. Ho modificato immagini e aggiunto la sbarra anche tra i geni della femmina di Drosophila.
ASSOCIAZIONE GENETICA parte 1
Come molti sapranno i cromosomi possiedono svariati geni, la domanda è: se i geni posti su cromosomi diversi assortiscono indipendentemente a causa della segregazione indipendente dei geni omologhi che si verifica durante la meiosi, in che modo assortiscono i geni sullo stesso cromosoma?
Tipico esempio: due geni legati al cromosoma X del moscerino della frutta che determinano il colore degli occhi e il colore del corpo.

Una femmina di Drosophila con occhi bianchi e corpo grigio wg+/wg+ (i geni li indicheremo rispettivamente come w= white; g+= gray cioè grigio) viene incrociata con maschi con occhi rossi e corpo viola w+g/Y (w+= gene selvatico; g= gene mutante), la progenie F1 è costituita da 50% di femmine con colore del corpo grigio e occhio rosso e dal 50% di maschi con occhio bianco e corpo grigio.
Da notare che i maschi presentano lo stesso fenotipo delle madri della generazione parentale.
Abbiamo detto che questi cromosomi sono presenti sul cromosoma X, i maschi possiedono un solo cromosoma X chè stato ereditato dalla madre. Discorso diverso per le femmine che hanno ricevuto un X dalla madre e un X dal padre; con due alleli per ciascuno dei cromosomi X, le dominanze fanno il resto.
Tale progenie viene incrociata e si ottiene una F2 che presenterà un totale di individui maschi di 7030, di cui 3600 presenteranno occhi bianchi e corpo grigio; 3360 presenteranno occhi rossi e corpo viola; mentre 80 individui presenteranno occhi rossi con corpo grigio e 50 individui presenteranno occhi bianchi con corpo viola.

Se i geni per il colore del corpo e dell'occhio assortissero in maniera indipendente come previsto dalla legge di Mendel le femmine F1 dovrebbero produrre quattro gameti con quattro combinazioni diverse dei geni sul cromosoma X cioè dovremmo avere gameti con queste combinazioni wg+; w+g; w+g+; wg.
Questi gameti dovrebbero essere presenti con la stessa frequenza e in un rapporto di 1:1:1:1.
Se così fosse allora approssimativamente metà dei gameti sarebbe di due tipi parentali e porterebbe o la combinazione allelica della madre o quella del padre (ci riferiamo a maschi e femmine della generazione P).
La metà restante sarebbe di due tipi ricombinanti, in cui il mescolamento ha prodotto le combinazioni alleliche w+g+; wg non presenti in nessuno degli individui della generazione P.
Si può veficare se si è effettivamente ottenuto il rapporto 1:1:1:1 nei gameti, contando il numero degli individui della generazione F2 esposta sopra, dal momento che ricevono alleli dal gene X che proviene solo dalla madre.
Come accennato sopra il numero totale degli individui è 7030 maschi, come possiamo notare abbiamo un significativo spostamento dal rapporto 1:1:1:1 che ci attenderemmo nel caso di assoritmento indipendente.
I gameti che portano combinazioni parentali: occhi bianchi/corpo marrone (wg+) e occhi rossi/corpo viola (w+g) sono tanti, di gran lunga superiori ad un rapporto di 1:1:1:1. Infatti il 99% presenta caratteri parentali ; le nuove combinazioni, quelle ricombinanti sono l'1% del totale.
Perchè avviene questo?
Una possibilità potrebbe essere che la combinazione parentale che le femmine F1 ricevono da uno o l'altro dei genitori si presenti per qualche motivo con più frequenza.
Una ulteriore serie di incroci ci può dare la risposta. Infatti come vedremo nel prossimo post, una preponderanza dei genotipi parentali nella generazione F2 definisce l'associazione.
Nota: l'immagine sopra quella dei due sgorbi che dovrebbero essere le teste della Drosophila, non chiedetemi perchè ma non ho resistito, le ho scopiazzate, intendo la forma, i colori dell'esempio li ho scelti io a caso, da un libro di genetica, Principi di genetica Snustad Simmons. Prometto che nei prossimi post cerco immagini più belle e smetto di fotografare ciò che scarabocchio sul foglio!!!
ASSOCIAZIONE, RICOMBINAZIONE E MAPPATURA GENICA
Un lettore mi aveva chiesto di parlare di questi argomenti, avrei dovuto pubblicarli tutti entro pomeriggio di oggi 14 novembre, purtroppo lo sciopero dei treni mi ha bloccato a Napoli e sono stato costretto ad un odissea che mi ha riportato a casa ad ora tarda, il post sulla associazione e ricombinazione non l'ho pubblicato in quanto non ho trovato immagini adatte, entro domattina, e stavolta MANTERRO' LA PAROLA saranno pubblicati, spero possano essere di aiuto! Nel frattempo sotto a questo post troverete l'argomento riguardante la mappatura genica.
MAPPATURA GENETICA: localizzazione dei geni sui cromosomi.
A.H.Sturtevant, studente di Morgan come argomento per la tesi di laurea si chiese se i dati ottenuti da un vasto numero di incroci a due punti (incroci che permettono di seguire due geni alla volta) potessero suffragare l'idea che i geni costituiscono una serie lineare su un cromosoma, cioè che siano disposti secondo uno schema lineare.
Esempio: consideriamo tre geni w,y, m. Se i geni sono disposti in una struttura lineare allora uno di essi deve trovarsi in una posizione centrale paragonata a quella degli altri due. La distanza genetica più grande quella che separa i geni più esterni e questo valore dovrebbe essere uguale alla somma delle distanze che separano il gene centrale da ciascuno dei geni laterali.
La mappatura per ricombinazione sembrava dare ragione all'ipotesi che i geni sono disposti su una struttura "lineare".
Sturtevant stabilì un ordine non contradditorio di tutti i geni da lui studiati sul cromosoma X, controllando i dati per ogni combinazione di tre geni, ci si rende conto che qusto ordine aveva una logica. Inoltre i dati sulla ricombinazione erano a favore della teoria che si potesse costruire una mappa in cui i geni erano disposti linearmente lungo il cromosoma.

 La mappatura però ha dei limiti, infatti negli incroci che coinvolgono solo due geni alla volta può essere difficile riuscire a trovare l'ordine esatto dei geni se i due geni sono particolarmente vicini tra di loro.
La mappatura però ha dei limiti, infatti negli incroci che coinvolgono solo due geni alla volta può essere difficile riuscire a trovare l'ordine esatto dei geni se i due geni sono particolarmente vicini tra di loro.
Per esempio, mappando y, m, w, la distanza tra i due geni esterni y e m è 34,3 u.m, la distanza tra i geni entrale w e l'esterno m è circa dello stesso ordine di grandezza 32,8 u.m.; come possiamo notare i valori sono molto simili, per evitare di trovarci difronte ad un errore di campionamento, bisognerebbe incrociare un grandissimo numero di moscerini e sottoporre i dati ad un test statisctico, come il test del chi quadrato.
Inoltre n altro problema che si incontra con il metodo di mappatura di Sturtevant è che le distanze effettive nella mappa non sono sempre addittive.
Esempio: se il locus del gene y si trova all'estrema sinistra della nostra appa n una posizione che possiamo definire come punto zero; il gene w sarebbesituato allora vicino alla posizione 1, ed m nelle vicinanze della posizione 34,2 u.m.
Per il gene r basandoci su quella che è la distanza da y (osserviamo la figura a) dovrebbe trovarsi in posizione 42,9 u.m. il problema sorge quando andiamo a sommare la distanza intermedia, cioè addizionando le distanze y<-->w più la distanza w<--->v più la distanza v<--->m più la distanza il locus r dovrebbe trovarsi ad una distanza di circa 55 u.m. Quale valore tra i due è più vero?
l'incrocio a tre punti può darci qualche risposta a riguardo
Incrocio a tre punti.
Di cosa si tratta questo tipo di incrocio? Nell'incrocio a tre punti analizziamo simultaneamente tre geni, i nostri marcatori, l'utilizzo dell'incrocio a tre punti ci permette di dAnalizzando simultaneamente tre marcatori, possiamo ottenere più facilmente una serie di informazioni che ci permettono di identificare l'esatta posizione dei geni.
Esempio: femmina omozigote per i caratteri ali vestigiali (a); corpo nero (b) occhi gialli (c) viene incrociata con maschio di tipo selvatico con geni a+ b+ c+.
La progenie F1 è eterozigote e mostra, sia per i maschi che per le femmine, un fenotipo normale per i tre caratteri indicando che le mutazioni sono autosomiche e recessive, la progenie F1 è rappresentata da individui tutti identici.
 Si esegue un testcross, nel quale incrociamo femmine eterozigoti della F1 e maschi selvatici; otteniamo una progenie che presenterà diversi fenotipi che rispecchiano ovviamente differenti genotipi.
Si esegue un testcross, nel quale incrociamo femmine eterozigoti della F1 e maschi selvatici; otteniamo una progenie che presenterà diversi fenotipi che rispecchiano ovviamente differenti genotipi.
Noi non conosciamo l'ordine dei geni, potrebbe essere a b c o qualsiasi altra combinazione derivante ed indicare lo stesso genotipo, cioè non conoscendo il modo in cui tali geni sono disposti sul cromosoma, noi potremmo anche scrivere b a c oppure c a b ed indicare lo stesso genotipo e fenotipo.
Lo scopo di mappare però è quello di riuscire a comprendere in che ordine sono disposti i geni di nostro interesse su un cromosoma.
 Osserviamo i dati della progenie del testcross nell'immagine sopra.
Osserviamo i dati della progenie del testcross nell'immagine sopra.
Ora prendiamo in considerazione due geni alla volta, numero questo fondamentale per eseguire il calcolo della frequenza di ricombinazione.
Prendiamo la coppia di geni a e b. Le combinazioni parentali sono a+ e b+; i ricombinanti sono a e b+; a+ e b.
Per determinare se una classe particolare della progenie è ricombinante o parentale per queste due classi di geni non importa sapere se i nostri moscerini sono c+ o c.
La distanza a<-->b calcolata come la percentuale di ricombinanti sul numero totale della progenie sarà di 350+338+231+218/5109 x 100= 22,5 u.m
 Lo stesso ragionamento che abbiamo applicato per a e b lo applichiamo ad a e c; osservando sempre i dati sopra abbiamo che:
Lo stesso ragionamento che abbiamo applicato per a e b lo applichiamo ad a e c; osservando sempre i dati sopra abbiamo che:
350+338+15+7/5109 x 100= 13,89 u.m
Stesso discorso anche per b e c:
231+218+15+7/5109 x 100= 9,3 u.m
Ciò dimostra che a e b sono la coppia di geni più distanti e di conseguenza possiamo assumere che sul cromosoma sono i geni presenti in posizione più esterna, con il gene c che è localizzato tra questi due geni, quindi in una posizione che potremmo definire "centrale".
Anche in questo caso però possiamo riscontrare lo stesso problema analizzato nell'incrocio a due punti; la distanza tra i geni più esterni non corrisponde alla somma delle distanze intermedie, infatti 13,89 + 9,3 = 23,20 u.m.
Il motivo? Dobbiamo considerare i doppi crossing over!
Prendiamo sempre in considerazione questi tre geni ,quelli che ho provato a disegnare, sono cromosomi autosomi omologhi eterozigoti per i geni di cui abbiamo accennato sopra.
Osservando l'immagine possiamo notare quali sono le regioni coinvolte nel crossing over che hanno dato origine alla progenie osservata precedentemente.
La progenie che avevamo ottenuto in un testcross tra femmine della F1 triple eterozigoti e maschi selvatici omozigoti per alleli recessivi di tutti e tre i caratteri è costituita da otto gruppi.
Ogni gruppo è costituito da un numero differente di moscerini; i due gruppi più numerosi hanno la stessa configurazione genica dei loro nonni, la generazione P (a b c e a+ b+ c+) questi geni costituiscono la classe parentale.
I gruppi successivi (a+ b c e a b+ c+) sono le classi ricombinanti che rappresentano i prodotti reciproci di un crossing over nella regione 1 tra a e c.
 Analogamente i due gruppi contenenti individui a+ b c+ e a b+c sono il risultato della ricombinazione avvenuta nella regione 2 tra c e b.
Analogamente i due gruppi contenenti individui a+ b c+ e a b+c sono il risultato della ricombinazione avvenuta nella regione 2 tra c e b.
 Arriviamo infine ai due gruppi meno numerosi, essi sono rari ricombinanti a b c+ e a+ b+ c.
Arriviamo infine ai due gruppi meno numerosi, essi sono rari ricombinanti a b c+ e a+ b+ c.
In questi due gruppi si sono verificati degli eventi di doppi crossing over differenti avvenuti contemporaneamente, uno nella regione 1 del cromosoma l'altro nella regione 2.
 I gameti prodotti da tale doppio crossing over mantengono sempre la configurazione parentale per i geni esterni a e b anche se sono avvenuti due scambi.
I gameti prodotti da tale doppio crossing over mantengono sempre la configurazione parentale per i geni esterni a e b anche se sono avvenuti due scambi.
Bisogna tenere conto di questo doppio scambio, infatti la distanza che abbiamo precedentemente calcolato non ne teneva conto , quindi bisogna correggere l'errore nel calcolo della frequenza di ricombinazione aggiungendo a tale calcolo i doppi crossing over, dal momento che ogni individuo che mostra un genotipo tipico del doppio crossing over è il risultato di due scambi tra a e b.
350+338+231+218+15+15+7+7/5109 x100= 23,16
questo valore ora risulterà più corretto, se andiamo a fare i conti e sommiamo anche le distanze intermedie tra i due geni ci renderemo conto che la distanza tra a e b è identica alla somma della distanza tra a e c e tra c e b.
Quando Sturtevant inizialmente mappò i geni mediante incroci a due punti, la posizione del locus per il gene delle ali rudimentali risultava ambigua. Un incrocio a due punti dava una frequenza di ricombinazione di 42,9 mentre la somma delle distanze intermedie dava un valore più elevato, 55,0.
Questo era dovuto al fatto che la mappa a due punti ignora il processo del doppio crossing over avvenuto nel vasto intervallo dei geni y e r. Invece sommando i valori di distanze più piccole ciò può essere notato più facilmente anche perchè un evento di doppio crossing over difficilmente avviene in intervalli molto piccoli.
Bisogna sottolineare però che anche una mappa a tre punti non prende in considerazione a priori il processo del doppio crossing over.
Quando si costruisce una mappa è meglio sempre utilizzare più geni possibili separati da distanze brevi.
Esempio: consideriamo tre geni w,y, m. Se i geni sono disposti in una struttura lineare allora uno di essi deve trovarsi in una posizione centrale paragonata a quella degli altri due. La distanza genetica più grande quella che separa i geni più esterni e questo valore dovrebbe essere uguale alla somma delle distanze che separano il gene centrale da ciascuno dei geni laterali.
La mappatura per ricombinazione sembrava dare ragione all'ipotesi che i geni sono disposti su una struttura "lineare".
Sturtevant stabilì un ordine non contradditorio di tutti i geni da lui studiati sul cromosoma X, controllando i dati per ogni combinazione di tre geni, ci si rende conto che qusto ordine aveva una logica. Inoltre i dati sulla ricombinazione erano a favore della teoria che si potesse costruire una mappa in cui i geni erano disposti linearmente lungo il cromosoma.

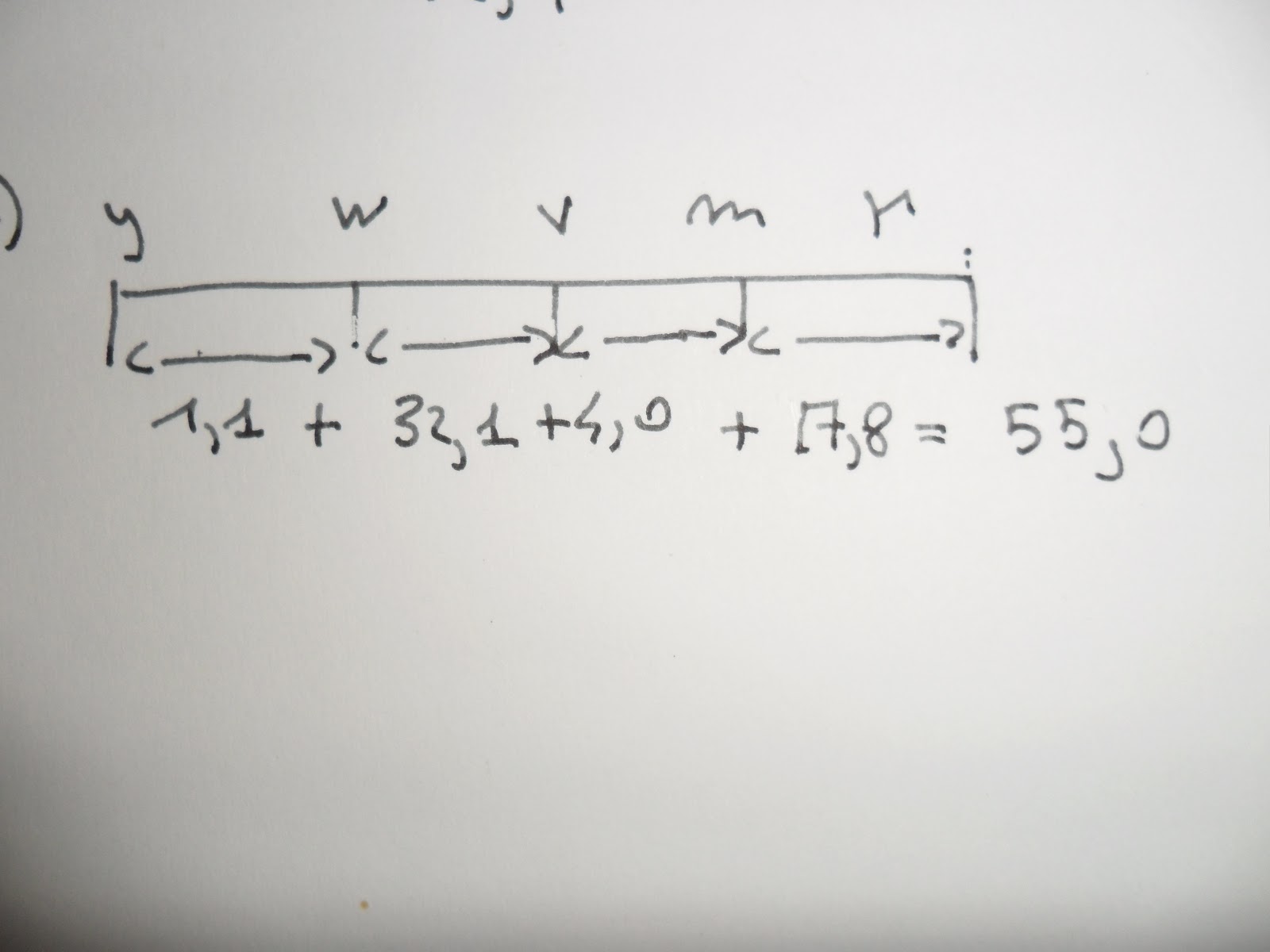
Per esempio, mappando y, m, w, la distanza tra i due geni esterni y e m è 34,3 u.m, la distanza tra i geni entrale w e l'esterno m è circa dello stesso ordine di grandezza 32,8 u.m.; come possiamo notare i valori sono molto simili, per evitare di trovarci difronte ad un errore di campionamento, bisognerebbe incrociare un grandissimo numero di moscerini e sottoporre i dati ad un test statisctico, come il test del chi quadrato.
Inoltre n altro problema che si incontra con il metodo di mappatura di Sturtevant è che le distanze effettive nella mappa non sono sempre addittive.
Esempio: se il locus del gene y si trova all'estrema sinistra della nostra appa n una posizione che possiamo definire come punto zero; il gene w sarebbesituato allora vicino alla posizione 1, ed m nelle vicinanze della posizione 34,2 u.m.
Per il gene r basandoci su quella che è la distanza da y (osserviamo la figura a) dovrebbe trovarsi in posizione 42,9 u.m. il problema sorge quando andiamo a sommare la distanza intermedia, cioè addizionando le distanze y<-->w più la distanza w<--->v più la distanza v<--->m più la distanza il locus r dovrebbe trovarsi ad una distanza di circa 55 u.m. Quale valore tra i due è più vero?
l'incrocio a tre punti può darci qualche risposta a riguardo
Incrocio a tre punti.
Di cosa si tratta questo tipo di incrocio? Nell'incrocio a tre punti analizziamo simultaneamente tre geni, i nostri marcatori, l'utilizzo dell'incrocio a tre punti ci permette di dAnalizzando simultaneamente tre marcatori, possiamo ottenere più facilmente una serie di informazioni che ci permettono di identificare l'esatta posizione dei geni.
Esempio: femmina omozigote per i caratteri ali vestigiali (a); corpo nero (b) occhi gialli (c) viene incrociata con maschio di tipo selvatico con geni a+ b+ c+.
La progenie F1 è eterozigote e mostra, sia per i maschi che per le femmine, un fenotipo normale per i tre caratteri indicando che le mutazioni sono autosomiche e recessive, la progenie F1 è rappresentata da individui tutti identici.

Noi non conosciamo l'ordine dei geni, potrebbe essere a b c o qualsiasi altra combinazione derivante ed indicare lo stesso genotipo, cioè non conoscendo il modo in cui tali geni sono disposti sul cromosoma, noi potremmo anche scrivere b a c oppure c a b ed indicare lo stesso genotipo e fenotipo.
Lo scopo di mappare però è quello di riuscire a comprendere in che ordine sono disposti i geni di nostro interesse su un cromosoma.

Ora prendiamo in considerazione due geni alla volta, numero questo fondamentale per eseguire il calcolo della frequenza di ricombinazione.
Prendiamo la coppia di geni a e b. Le combinazioni parentali sono a+ e b+; i ricombinanti sono a e b+; a+ e b.
Per determinare se una classe particolare della progenie è ricombinante o parentale per queste due classi di geni non importa sapere se i nostri moscerini sono c+ o c.
La distanza a<-->b calcolata come la percentuale di ricombinanti sul numero totale della progenie sarà di 350+338+231+218/5109 x 100= 22,5 u.m

350+338+15+7/5109 x 100= 13,89 u.m
Stesso discorso anche per b e c:
231+218+15+7/5109 x 100= 9,3 u.m
Ciò dimostra che a e b sono la coppia di geni più distanti e di conseguenza possiamo assumere che sul cromosoma sono i geni presenti in posizione più esterna, con il gene c che è localizzato tra questi due geni, quindi in una posizione che potremmo definire "centrale".
Anche in questo caso però possiamo riscontrare lo stesso problema analizzato nell'incrocio a due punti; la distanza tra i geni più esterni non corrisponde alla somma delle distanze intermedie, infatti 13,89 + 9,3 = 23,20 u.m.
Il motivo? Dobbiamo considerare i doppi crossing over!
Prendiamo sempre in considerazione questi tre geni ,quelli che ho provato a disegnare, sono cromosomi autosomi omologhi eterozigoti per i geni di cui abbiamo accennato sopra.
Osservando l'immagine possiamo notare quali sono le regioni coinvolte nel crossing over che hanno dato origine alla progenie osservata precedentemente.
La progenie che avevamo ottenuto in un testcross tra femmine della F1 triple eterozigoti e maschi selvatici omozigoti per alleli recessivi di tutti e tre i caratteri è costituita da otto gruppi.
Ogni gruppo è costituito da un numero differente di moscerini; i due gruppi più numerosi hanno la stessa configurazione genica dei loro nonni, la generazione P (a b c e a+ b+ c+) questi geni costituiscono la classe parentale.
I gruppi successivi (a+ b c e a b+ c+) sono le classi ricombinanti che rappresentano i prodotti reciproci di un crossing over nella regione 1 tra a e c.


In questi due gruppi si sono verificati degli eventi di doppi crossing over differenti avvenuti contemporaneamente, uno nella regione 1 del cromosoma l'altro nella regione 2.

Bisogna tenere conto di questo doppio scambio, infatti la distanza che abbiamo precedentemente calcolato non ne teneva conto , quindi bisogna correggere l'errore nel calcolo della frequenza di ricombinazione aggiungendo a tale calcolo i doppi crossing over, dal momento che ogni individuo che mostra un genotipo tipico del doppio crossing over è il risultato di due scambi tra a e b.
350+338+231+218+15+15+7+7/5109 x100= 23,16
questo valore ora risulterà più corretto, se andiamo a fare i conti e sommiamo anche le distanze intermedie tra i due geni ci renderemo conto che la distanza tra a e b è identica alla somma della distanza tra a e c e tra c e b.
Quando Sturtevant inizialmente mappò i geni mediante incroci a due punti, la posizione del locus per il gene delle ali rudimentali risultava ambigua. Un incrocio a due punti dava una frequenza di ricombinazione di 42,9 mentre la somma delle distanze intermedie dava un valore più elevato, 55,0.
Questo era dovuto al fatto che la mappa a due punti ignora il processo del doppio crossing over avvenuto nel vasto intervallo dei geni y e r. Invece sommando i valori di distanze più piccole ciò può essere notato più facilmente anche perchè un evento di doppio crossing over difficilmente avviene in intervalli molto piccoli.
Bisogna sottolineare però che anche una mappa a tre punti non prende in considerazione a priori il processo del doppio crossing over.
Quando si costruisce una mappa è meglio sempre utilizzare più geni possibili separati da distanze brevi.
sabato 10 novembre 2012
L'ANALISI DEI RARI ERRORI MEIOTICI COME SOSTEGNO ALLA TEORIA CROMOSOMICA DELL'EREDITARIETA'.
Come abbiamo potuto vedere nel post precedente Morgan suppose che la mutazione per il colore degli occhi della Drosophila risiedesse sul cromosoma X, per dimostrarlo portò avanti vari incroci, bisogna dire però che Morgan non era comunque assolutissimamente convinto della teoria cromosomica dell'ereditarietà fino a quando Calvin Bridges, un suo studente, tramite vari studi ottenne un ulteriore prova chiave a carico della teoria cromosomica.
Bridges non fece che ripetere in maniera più estesa gli stessi esperimenti effettuati da Morgan, incrociando femmine con occhio bianco e maschi con occhi rossi.
Come ci si aspettava la progenie era costituita da femmine con occhi rossi e maschi con occhi bianchi.
Notò però che 1 su 2000 aveva occhi rossi e con la stessa percentuale, più o meno si avevano femmine con occhi bianchi.


Queste osservazioni incuriosirono Bridges il quale inizio a supporre che queste eccezioni derivassero da rari eventi in cui i cromosomi X non riuscivano a separarsi durante la meiosi femminile.
Questa mancata segregazione venne definita "non disgiunzione".
Questa mancata segregazione venne definita "non disgiunzione".
Come mostrato nell'immagine sotto in conseguenza della non disgiunzione si otterrebbero alcune cellule uovo con due cromosomi X e altre senza cromosomi X.

Osservando i cromosomi della femmine con occhi bianchi ottenute da incroci su vasta scala, Bridges trovò che erano individui XXY che avevano ricevuto i due cromosomi X e con essi l'allele w dalle madri con occhio bianco.
I maschi eccezzionali con occhio rosso nati dall'incrocio erano XO, il colore dell'occhio mostrava che avevano ricevuto l'unico cromosoma sessuale dal padre con occhi rossi. Lo studio mise in evidenza che si erano verificati degli errori nella distribuzione dei cromosomi durante la divisione cellulare.
I moscerini OY non sopravvivono in quanto il cromosoma X porta geni necessari per la vitalità che non sono presenti sul cromosoma Y anche le femmine XXX morivano.
Ciò che possiamo notare è che le femmine con occhio bianco possiedono 3 cromosomi sessuali XXY, Bridges ipotizzò che potessero produrre quattro tipi di uova, XY e X oppure XX e Y.
La formazione di queste quattro uova può essere visualizzata se immaginiamo che i 3 cromosomi si appaiano e si dividono durante la meiosi, due cromosomi devono andare ad un polo e un cromosoma all'altro.
La formazione di queste quattro uova può essere visualizzata se immaginiamo che i 3 cromosomi si appaiano e si dividono durante la meiosi, due cromosomi devono andare ad un polo e un cromosoma all'altro.
Con questo tipo di segregazione i risultati possibili sono due: il cromosoma X e Y vanno insieme al secondo cromosoma X ad un polo, oppure i cromsomi X ad un polo e il cromosoma Y all'altro.
Quest'ultimo caso avviene raramente quando i cromosomi X non riescono ad appaiarsi.
Bridges previde che la fecondazione di queste quattro tipi di uova, da parte di spermatozooi normali avrebbe generato dei cariotipi per i cromosomi sessuali di un vasto numero di individui per la progenie.
 Infatti riuscì a dimostrare citologicamente (vedere immagine a lato e sotto) che le femmine con occhio bianco ottenute dall'incrocio con maschio occhi rossi, avevano due cromosomi X e un cromosoma Y, mentre metà dei maschi con occhio bianco mostrava un unico cromosoma X e due Y.
Infatti riuscì a dimostrare citologicamente (vedere immagine a lato e sotto) che le femmine con occhio bianco ottenute dall'incrocio con maschio occhi rossi, avevano due cromosomi X e un cromosoma Y, mentre metà dei maschi con occhio bianco mostrava un unico cromosoma X e due Y.
Ciò fornì le prove dell'ipotesi che i geni specifici sono realmente localizzati su cromosomi specifici.

mercoledì 7 novembre 2012
MITOSI E MEIOSI A CONFRONTO.
La mitosi.
La mitosi è il principale processo che assicura che ogni cellula di un organismo porti gli stessi cromosomi, mantenendo il numero corretto di cromosomi in ogni cellula.
 Interfase.
Interfase.

Il DNA è contenuto nel nucleo delle cellule, quando la cellula non è in fase di divisione cellulare (interfase) il nucleo è intatto, I cromosomi durante questa fase appaiono come aggregati indistinguibili di cromatina.
 Durante la profase l'inizio della mitosi è rappresentato da un condensamento dei singoli cromosomi, processo appunto noto come condensazione. Ogni cromosoma che si condensa ha già subito il processo della replicazione ed è costituito da due cromatidi fratelli uniti tra di loro da una struttura nota come centromero.
Il processo di condensazione è straordinario, la macromolecola del DNA come molti sapranno è molto grande, potrebbe raggiungere una lungezza di svariati centimetri, per essere contenuta nel nucleo deve essere letteralmente impacchettata attraverso complessi e specifici processi.
Durante la profase l'inizio della mitosi è rappresentato da un condensamento dei singoli cromosomi, processo appunto noto come condensazione. Ogni cromosoma che si condensa ha già subito il processo della replicazione ed è costituito da due cromatidi fratelli uniti tra di loro da una struttura nota come centromero.
Il processo di condensazione è straordinario, la macromolecola del DNA come molti sapranno è molto grande, potrebbe raggiungere una lungezza di svariati centimetri, per essere contenuta nel nucleo deve essere letteralmente impacchettata attraverso complessi e specifici processi.
Con la condensazione i cromosomi si trasformano da uno stat metabolicamente attivo ad una cndizione che li rende pronti per ilsucessivo trasporto nelle cellule figlie.
Durante questa fase i nucleoli iniziano a scomparire. I centrosomi si sono duplicati in interfase, si dividono e sono chiaramente distinguibili come due entità separate visibili al microscopio ottico. Nello stesso tempo una sottile e fitta rete di microtubuli dell'interfase si disaggrega ed è sostituita da microtubuli che crescono e si contraggono rapidamente verso i propri centri di organizzazione dei centrosomi. I centrosomi continuano a separarsi migrano attorno alla membrana nucleare verso le estremità opposte del nucleo.
 I cinetocori contengono proteine che agiscono da motori molecolari, permettendo ai cromosomi di scivolare lungo i microtubuli.
I cinetocori contengono proteine che agiscono da motori molecolari, permettendo ai cromosomi di scivolare lungo i microtubuli.

 Metafase: Il collegamento dei cromatidi fratelli innesca una serie di reazioni che permettono ai cromosomi di muoversi verso l'equatore immaginario, la cosidetta piastra metafasica.
Metafase: Il collegamento dei cromatidi fratelli innesca una serie di reazioni che permettono ai cromosomi di muoversi verso l'equatore immaginario, la cosidetta piastra metafasica.
Quando i cromosomi si allineano su di essa le forze che tirano i cromatidi verso i poli e le forze che lo mantengono in una posizione equidistante dai poli si equivalgono.
Anafase: durante l'anafase avviene la separazione dei cromosomi fratelli, ciò fa in modo che ogni cromatidio venga tirato verso il polo a cui è collegato tramite i microtubuli del cinetocore non appena il cromosoma si muove verso il polo, i microtubuli del cinetocore si accorciano.
Dato che le braccia dei cromatidi metacentrici seguono passivamente i cinetocori, assumono una configurazione caratteristica a forma di V durante l'anafase.
La connessione dei cromatidi fratelli ai microtubuli emanati dai poli opposti del fuso sta a significare che l'informazione genetica che migra verso un polo è esattamente uguale a quella che muove verso il polo opposto.
Telofase: la trasformazione finale del nucleo durante la mitosi avviene in telofase. La telofase è come la profase al contrario, le fibre del fuso iniziano a disperdersi e a livello di ciascun polo si riforma una membrana nucleare intorno al gruppo di cromatidi e ricompaiono i nucleoli. I cromatidi ora si comportano come cromosomi indipendenti che si decondensano e si despiralizzano fino a costituire un massa affrovigliata di cromatina. La mitosi finisce qui.
La meiosi e la produzione di cellule germinali aploidi, i gameti.
Durante i vari cicli di divisioni embrionali, la maggior parte delle cellule possono crescere e dividersi attraverso la mitosi o arrestarsi in G0.
Le cellule che vanno in divisione mitotica e quelle arrestate nella fase G0 sono chiamate cellule somatiche, e le cellule che derivano da esse andranno a costituire gran parte dei tessuti di un organismo durante il suo ciclo vitale. Altre cellule avranno un destino del tutto diverso; sono le cellule germinali; il loro destino sarà quello di specializzarsi nella produzione dei gameti. Le cellule germinali faranno in seguito parte degli organi riproduttivi (testicoli, ovaie negli animali, fiori e antere nelle piante) dove infine andranno incontro alla meiosi, la divisione a due fasi che produce i gameti (cellule uova e spermatozoi o polline).
Tali cellule conterranno metà numero di cromosomi rispetto alle altre cellule dell'organismo.
Le cellule somatiche possiedono vita limitata; tendono ad andare incontro ad un certo numero di divisioni cellulari mitotiche e poi muoiono. Al contrario quelle germinali tendono ad acquistare una sorta di immortalità in quanto, generando gameti, trasmettono i geni di un indviduo alla generazione successiva.
La meiosi è un processo cellulare mediante il quale il materiale cromosomico si replica una volta mentre la cellula si divide due volte, questo meccanismo ha l'importante funzione di diminuire il numero di cromosomi all'interno delle cellule figlie. Nelle cellule pre-meiotiche ogni cromosoma è appaiato con un omologo, e per ogni coppia di omologhi un cromosoma è dato dallo spermatozoo (contributo paterno) e un cromosoma è dato dall'uovo (contributo materno). Alla conclusione della meiosi ogni cellula sarà aploide, cioè mostrerà solo un membro della coppia di cromosomi omologhi sia esso materno o paterno. La meiosi nei primi stadi dello sviluppo di un organismo è di fondamentale importanza in quanto assicura una costanza nel numero cromosomico attraverso le generazioni di una particolare specie. Infatti se ciò non avvenisse la fusione di due cellule dipolidi produrrebbe uno zigote con il doppio dei cromosomi presenti nella generazione parentale, ad esempio nell'uomo il numero dei cromosomi in una cellula diploide è 46, la fusione di due cellule diploidi produrrebbe in seguito uno zigote con 92 cromosomi, nelle generazione successiva il doppio!
Inoltre errori nella segregazione dei cromosomi durante le fasi meiotiche possono portare a delle anomalie a livello genetico, come il processo definito di non disgiunzione nel quale i cromosomi non segregano correttamente portando alla formazione di gameti che presentano un numero maggiore di cromosomi in un gamete rispetto al normale e in un altro gamete essere assenti, oppure essere presenti in un numero deficitario rispetto al normale. La stragrande maggioranza di queste anomalie sono letali sia che coinvolgano cromosomi autosomici o sessuali, in alcuni casi come la trisomia 21 che causa la sindrome di down non è mortale ma porta modificazioni fenotipiche che possono essere molto debilitanti per l'individuo che ne è affetto.
In altri casi ancora a differenza dei casi in cui si verificano errori nella segregazione dei cromosomi, alcuni animali ibridi portano cromosomi non omologhi che non possono segregare correttamente. Un esempio è l'unione tra il cavallo e l'asino che produce il mulo che come molti sanno è sterile. Ciò è dovuto al fatto che il padre asino porta 30 cromosomi il cavallo 32 e molti di questi cromosomi sono troppo differenti per potersi appaiare, cio provoca formazione di gameti non vitali nel mulo con conseguente sterilità.
Abbiamo accennato al fatto che la meiosi svolge un ruolo di fondamentale importanza nell'assicurare la costanza del numero cromosomico nelle generazioni successive, ma allo stesso tempo svolge un ruolo primario nell'assicurare la variabilità genetica nelle generazioni. Infatti la meiosi presenta due aspetti di fondamentale importanza nell'assicurare la variabilità genetica. Primo è del tutto casuale quale dei cromosomi migri verso un determinato polo della cellula, 2) gameti diversi portano combinazioni genetiche differenti. Inoltre dobbiamo tenere conto anche del fatto che più aumenta l'assortimento di combinazioni geniche in una popolazione maggiori saranno le possibilità che alcuni individui possiedano combinazioni di alleli che possono permettere la sopravvivenza di un individuo in un particolare ambiente, e inoltra maggiore sarà il numero di cromosomi maggiore la variabilità genetica. Ad esempio negli esseri umani il numero di cromosomi in un gamete è 23 l'assortimento casuale di gameti con i rispettivi cromosomi potrebbe produrre 2 elevato alla 23 tipi di gameti differenti...(quindi 2 elevata alla 23 combinazioni genotipiche differenti).
Una seconda caratteristica poi è il mescolamento dei cromosomi che avviene durante il processo del crossing over in cui i cromosomi scambiano tra di loro segmenti di materiale genetico, contribuendo alla variabilità genetica, senza contare che la riproduzione sessuale amplifica questa variabilità, durante la riproduzione di uno qualsiasi dei tanti spermatozooi geneticamente diversi può fecondare uno dei tanti ovuli con la propria distinta identità genetica.
La meiosi è costituita da due cicli di divisione cellulare.
Caratteristica principale della meiosi è che a differenza della mitosi avengono due divisioni nucleari anzichè una sola.
Nella meiosi I l'evento principale è costituito dall'appaiamento dei cromosomi omologhi, dallo scambio di materiale genetico tra i cromosomi e infine dalla loro separazione.

Profase I: è la fase più lunga del processo meiotico, è il momento in cui avviene il crossing over.
La profase può essere suddivisa in vari sottostadi, il primo è noto come leptotene; all'avvio della meiosi I il dna è stato già duplicato, è la fsi in cui i cromosomi iniziano ad ispessirsi. Tutti i cromosomi sono già stati duplicati all'inizio della meiosi I ed ognuno è costituito da un cromatidio fratello attaccati d una struttura nota come centromero. In questo stadio i cromatidi sono strettamente associati tra di loro da apparire come un unica identità. Il successivo stadio è noto come zigotene, inizia questo stadio quando tutti i cromosomi omologhi sono appaiati tra di loro e inizian ad unirsi come una iusura lampo, a formare la sinapsi. QUesto processo avviene grazie all'azione e alla formazione di una elaborata e complessa struttura proteica nota come complesso sinaptinemale, è importante la formazione di tale complesso affinchè l'appaiamento dei cromosomi avvenga con estrema precisione, giustapponendo le stesse regioni genetiche dei due cromosomi.
Il pachitene; è lo stadio in cui avviene il crossing over, i cromosomi sono ora appaiati per tutta la loro lunghezza, i cromosomi X e Y sono differenti; l'appaiamento nel loro caso avviene solo in prossimità di particolari zone omolghe che permettono un appaiamento limitato. Durante il pachitene iniziano a formarsi delle particolari strutture denominate noduli di ricombinazione. A livello dei noduli di ricombinazione avviene uno scambio di materiale genomico tra i cromatidi non fratelli (materni e paterni). Lo scambio noto come crossing over, ha come risultato la ricombinazione di materiale genetico. Come risultato ogni cromosoma della coppia potrebbe non essere più di derivazione paterna e materna, ognuno comunque ha ancora un entità genomica completa dal momento che non vi è interruzione nell'informazione genetica.
 Il diplotene ha inizio ha inizio con il dissolvimento del complesso sinaptinemale e con una leggera separazione dei cromosomi omologhi. I cromosomi omologhi allinearti di ciascun cromosoma rimangono legati ancora attraverso particolari strutture note come chiasmi, che sembrano essere i luoghi dove è avvenuto il crossing over. La diacinesi è accompagnata da una condensazione ulteriore dei cromatidi. A causa di questo ispessimento e accorciamento dei cromosomi può notare che ogni tetrade è costituita da quattro cromatidi fratelli tenuti insieme da un centromero. I cromatidi fratelli che sono andati incontro al crossing-over restano associati strettamente all'altezza dei chiamsi. Il completamento della diacinesi è analogo a quello della prometafase mitotica: la membrana nucleare si disaggrega e iniziano a comparire le prime fibre del fuso.
Il diplotene ha inizio ha inizio con il dissolvimento del complesso sinaptinemale e con una leggera separazione dei cromosomi omologhi. I cromosomi omologhi allinearti di ciascun cromosoma rimangono legati ancora attraverso particolari strutture note come chiasmi, che sembrano essere i luoghi dove è avvenuto il crossing over. La diacinesi è accompagnata da una condensazione ulteriore dei cromatidi. A causa di questo ispessimento e accorciamento dei cromosomi può notare che ogni tetrade è costituita da quattro cromatidi fratelli tenuti insieme da un centromero. I cromatidi fratelli che sono andati incontro al crossing-over restano associati strettamente all'altezza dei chiamsi. Il completamento della diacinesi è analogo a quello della prometafase mitotica: la membrana nucleare si disaggrega e iniziano a comparire le prime fibre del fuso.
Metafase I: la differenza tra il fuso che si viene a formare durante questo processo e quello che si forma nella metafase mitotica è che durante la meiosi ogni cromatidio fratello possiede un cinetocore in grado di poter interaginre con i microtubuli che originano dai poli della opposti al fuso.
Inoltre in meiosi I i cinetocori dei cromatidi fratelli vanno incontro a fusione, ciò farà si che ogni cromosoma contenga un solo cinetocore.
La presenza di un solo cinetocore ha una conseguenza, durante l'anafase non vi sono forze opposte in grado di tirare i cromatidi fratelli da un lato e dall'atro. Nella metafase I al contrario le strutture dei microtubulu provenienti da poli opposti si attaccano ai cinetocori dei cromosomi omologhi.
I cromosomi in questo modo sono allineati nella piastra metafasica, i cinetocori dei cromosoi materni e paterni si affaceranno ai poli opposti del fuso, posizionando i cromosomi omologhi in modo tale che durante l'anafase possano venire tirati in direzioni opposte.
Dal momento che l'allineamento e l'aggancio di ciascun bivalente alle fibre del fuso sono indipendenti da quelli di qualsiasi altro bivalente, i cromosomi che si affacceranno ad un polo saranno un miscuglio casuale di cromosomi di origine materna e paterna.
Anafase I: al momento dell'anafase I i chiasmi che uniscono i cromosomi omologhi si dissolvono, permettendo agli omologhi materni e paterni di muoversi verso i poli del fuso.
Nella prima divisione meiotica i centromeri non si dividono come in meiosi; così per ogni bivalente migregà verso ciascun polo solo un cromosoma, costituito da due cromatidi fratelli uniti al centromero.
Durante questa fase avviene la ricombinazione tramite il processo definito del crossing over, il quale riveste un ruolo di fondamentale importanza nella segregzione dei cromosomi omologhi durante la prima divisione meiotica.
Questo perchè i chiasmi, mantenendo uniti i cromosomi omologhi, assicurano anche che i centromeri rimangano attaccati ai poli opposti del fuso per tutta la durata della metafase.
Quando in un bivalente la ricombinazione non avviene si possono verificare dei grvi errori nell'aggncio, e nel trasporto dei cromosomi, che possono portare ad un movimento dei cromosomi omologhi verso lo stesso polo invece che tra i poli opposti.
Telofase I: Durante il processo della telofase iniziano a formarsi le membrane nucleari che si formano attorno ai cromosomi che sono migrati ai poli.
I nuclei figli che si stanno per formare contengono ciascuno metà dei cromosomi del nucleo parentale, ma ciascun cromosoma è costituito da cromatidi fratelli uniti al centromero, durante la meiosi il numero di cromosomi si riduce alla metà del normale numero diploide (quindi si ha in questo momento un corredo aploide).
 Durante la meiosi II, i cromatidi fratelli si separano e si ha la produzione dei gameti aploidi.
Durante la meiosi II, i cromatidi fratelli si separano e si ha la produzione dei gameti aploidi.
Le fasi che caraterizzano la meiosi II sono più o meno identiche alla meiosi I:
Profase II: Nell'interfase precedente alla meiosi I i cromosomi si sono decondensati, ora si ricondensano al termine della profase II la membrana nucleare si dissolve nuovamente e si riforma il fuso.
Metafase II: durante tale fase i cinetocori dei cromatidi fratelli si attaccano al fibre del fuso, come nella metafase mitotica.
Tale fase però possiede una caratteristica fondamentale che la distingue da quella mitotica. Il numero dei cromosomi è la metà di quello presente nella metafase mitotica e nella maggior parte dei cromosomi i due cromatidi fratelli non sono più perfettamente identici a causa della ricombinazione avvenuta durante la meiosi I, i cromatidi possiedono ancora gli stessi geni ma possono portare combinazioni differenti di alleli.
Anafase II: come accade nella mitosi anche in questa fase i centromeri si separano e ciò permette ai cromatidi di migrare verso i poli opposti.
Telofase II: si sono formate quatttro cellule figlie si riformano le membrane e la citochinesi colloca i nuclei in cellula distinte. Il risultato è la formazione di quattro gameti aploidi .Alla fine di questa fase ognuno dei gameti ha lo stesso numero di cromosomi della cellula da cui si origina. La meiosi due per tale motivo viene definito divisione equazionale.
La mitosi è il principale processo che assicura che ogni cellula di un organismo porti gli stessi cromosomi, mantenendo il numero corretto di cromosomi in ogni cellula.
Il tipo di divisione cellulare che ripartisce i cromosomi in maniera equivalente tra le cellule figlie, il meccanismo cellulare che porta alla conservazione del materale genetico attraverso le varie generazioni cellulari. Quando si parla di mitosi ci si riferisce al processo finale di un ciclo cellulare in cui si assiste alla seprarazione di una cellula madre in due cellule figlie, infatti esso rappresenta solo una piccola parte del ciclo cellulare, il periodo che intercorre tra di essa è nota come interfase.
Dunque con il termine mitosi intendiamo il processo di divisione cellulare, solo una piccola parte del ciclo cellulare è utilizzato per la divisione della cellula. Il periodo che intercorre tra le divisioni è noto come interfase.
L'interfase a sua volta è suddivisibile in tre stadi che vengono rispettivamente chiamati G1-S-G2.
La fase G1 parte dalla nascita di una nuova cellula, a sua volta derivata da un precedente processo di mitosi.
Durante la G1 la cellula utilizza l'informazione genetica che ha ereditato per portare avanti tutte le sue funzioni. Questa fase comunemente è nota come di accrescimento, è molto variabile anche a seconda del tipo di cellula, per fare un esempio le cellule embrionali tendono a dividersi molto velocemente, le cellule del cervello invece tendono a sostare in una fase nota come fase G0, una fase di riposo della G1 e permangono in tale stato per tutta la durata della vita.
La mitosi la possiamo suddividere per comodità in vari stadi, ognuno dei quali caratterizzato da uno o più eventi cruciali. Le fasi della mitosi sono: profase, prometafase, metafase, anafase, telofase, citochinesi.
Come possiamo notare dall'immagine sopra, prima dell'inizio della fase G2 la cellula entra in fase S. Durante tale fase il materiale genetico viene duplicato.

Durante la replicazione, ciascun cromosoma si duplia per produrre cromatidi fratelli identici, che saranno visibili quando i cromosomi si condenseranno all'inizio della mitosi. I due cromatidi fratelli rimanfono uniti al centromero.
L'intervallo G2 è compreso tra la duplicazione dei cromosomi e l'inizio della mitosi. Durante questa fase la cellula subisce un accrescimento, anche se limitato rispetto alla fase G1 e sintetizzano le proteine fondamentali per i ossimi stadi cellulari. All'esterno del nucleo si può osservare l'inizio della formazione dei microtubuli che saranno fondamentali per le prossime fasi cellulari.
I microtubuli si irradiano all'interno del citoplasma a partire da un centro singolo di organizzazione noto con il termine di centrosoma, localizzato vicino alla membrana nucleare.
Nelle cellula animali il centrosoma è costituito nella sua parte centrale da due organelli scuri noti con il termine di centrioli . Durante la fase S e G2 i centrosomi si replicano producendo due centrosomi che rimangono molto vicini.
Profase: condensazione dei cromosomi.
 Durante la profase l'inizio della mitosi è rappresentato da un condensamento dei singoli cromosomi, processo appunto noto come condensazione. Ogni cromosoma che si condensa ha già subito il processo della replicazione ed è costituito da due cromatidi fratelli uniti tra di loro da una struttura nota come centromero.
Durante la profase l'inizio della mitosi è rappresentato da un condensamento dei singoli cromosomi, processo appunto noto come condensazione. Ogni cromosoma che si condensa ha già subito il processo della replicazione ed è costituito da due cromatidi fratelli uniti tra di loro da una struttura nota come centromero.Con la condensazione i cromosomi si trasformano da uno stat metabolicamente attivo ad una cndizione che li rende pronti per ilsucessivo trasporto nelle cellule figlie.
Durante questa fase i nucleoli iniziano a scomparire. I centrosomi si sono duplicati in interfase, si dividono e sono chiaramente distinguibili come due entità separate visibili al microscopio ottico. Nello stesso tempo una sottile e fitta rete di microtubuli dell'interfase si disaggrega ed è sostituita da microtubuli che crescono e si contraggono rapidamente verso i propri centri di organizzazione dei centrosomi. I centrosomi continuano a separarsi migrano attorno alla membrana nucleare verso le estremità opposte del nucleo.
Prometafase: La prometafase, inizia con la disgregazione della membrana nucleare, ciò permette ai microtubuli irradiati dai centrosomi di invadere la regione corrispondente al nucleo.
I cromosomi si attaccano ai microtubli mediante il cinetocore. I cinetocori contengono proteine che agiscono da motori molecolari, permettendo ai cromosomi di scivolare lungo i microtubuli.
I cinetocori contengono proteine che agiscono da motori molecolari, permettendo ai cromosomi di scivolare lungo i microtubuli.
Quando il cinetocore di un cromosoma inizialmente si attacca al microtubulo in prometafase, il motore che risiede sul cinetocore muove il cromosoma verso il centrosoma da cui quel microtubulo origina.
I microtubuli che crescono da due centrosomi catturano a caso i cromosomi attraverso i cinetocori di uno dei due cromatidi fratelli. Il risultato è che qualche volta è possibile osservare cromosomi che si raggruppano in prossimità di ciascun centrosoma.
In questa parte iniziale della prometafse un cromatidio per ogni cromosoma è attaccato a un microtubulo ma il cinetocore del cromatidio fratello rimane libero.

Il fuso mitotico è costituito da tre diversi tipi di fibre microtubulari: tutti questi microtubuli si originano dai centrosomi che funzionano come i due poli dell'apparato del fuso.
I microtubuli che si estendono tra un centrosoma e il cinetocore di un cromatidio sono i microtubuli del cinetocore oppure le fibre centromeriche.
I microtubuli che da ogni centrosoma sono diretti verso il centro della cellula sono definiti microtubuli polari; i microtubuli polari che si originano dai centrosomi opposti si interdigitano vicino l'equatore della cellula.
Ci sono infine, i corti microtubuli astrali, fibre poco stabili che si estendono dal centrosoma verso la periferia della cellula.
Verso la fine della prometafase i cinetocori dei cromatidi fratelli non ancora attaccati si associano ai microtubuli emanati dai centrosomi opposti.
In questo modo i cromosomi sono orientati in modo tale che un cromatidio fratello si affaccia verso un polo della cellula e l'altro verso il polo opposto.

Quando i cromosomi si allineano su di essa le forze che tirano i cromatidi verso i poli e le forze che lo mantengono in una posizione equidistante dai poli si equivalgono.
Anafase: durante l'anafase avviene la separazione dei cromosomi fratelli, ciò fa in modo che ogni cromatidio venga tirato verso il polo a cui è collegato tramite i microtubuli del cinetocore non appena il cromosoma si muove verso il polo, i microtubuli del cinetocore si accorciano.
Dato che le braccia dei cromatidi metacentrici seguono passivamente i cinetocori, assumono una configurazione caratteristica a forma di V durante l'anafase.
La connessione dei cromatidi fratelli ai microtubuli emanati dai poli opposti del fuso sta a significare che l'informazione genetica che migra verso un polo è esattamente uguale a quella che muove verso il polo opposto.
Telofase: la trasformazione finale del nucleo durante la mitosi avviene in telofase. La telofase è come la profase al contrario, le fibre del fuso iniziano a disperdersi e a livello di ciascun polo si riforma una membrana nucleare intorno al gruppo di cromatidi e ricompaiono i nucleoli. I cromatidi ora si comportano come cromosomi indipendenti che si decondensano e si despiralizzano fino a costituire un massa affrovigliata di cromatina. La mitosi finisce qui.
La meiosi e la produzione di cellule germinali aploidi, i gameti.
Durante i vari cicli di divisioni embrionali, la maggior parte delle cellule possono crescere e dividersi attraverso la mitosi o arrestarsi in G0.
Le cellule che vanno in divisione mitotica e quelle arrestate nella fase G0 sono chiamate cellule somatiche, e le cellule che derivano da esse andranno a costituire gran parte dei tessuti di un organismo durante il suo ciclo vitale. Altre cellule avranno un destino del tutto diverso; sono le cellule germinali; il loro destino sarà quello di specializzarsi nella produzione dei gameti. Le cellule germinali faranno in seguito parte degli organi riproduttivi (testicoli, ovaie negli animali, fiori e antere nelle piante) dove infine andranno incontro alla meiosi, la divisione a due fasi che produce i gameti (cellule uova e spermatozoi o polline).
Tali cellule conterranno metà numero di cromosomi rispetto alle altre cellule dell'organismo.
Le cellule somatiche possiedono vita limitata; tendono ad andare incontro ad un certo numero di divisioni cellulari mitotiche e poi muoiono. Al contrario quelle germinali tendono ad acquistare una sorta di immortalità in quanto, generando gameti, trasmettono i geni di un indviduo alla generazione successiva.
La meiosi è un processo cellulare mediante il quale il materiale cromosomico si replica una volta mentre la cellula si divide due volte, questo meccanismo ha l'importante funzione di diminuire il numero di cromosomi all'interno delle cellule figlie. Nelle cellule pre-meiotiche ogni cromosoma è appaiato con un omologo, e per ogni coppia di omologhi un cromosoma è dato dallo spermatozoo (contributo paterno) e un cromosoma è dato dall'uovo (contributo materno). Alla conclusione della meiosi ogni cellula sarà aploide, cioè mostrerà solo un membro della coppia di cromosomi omologhi sia esso materno o paterno. La meiosi nei primi stadi dello sviluppo di un organismo è di fondamentale importanza in quanto assicura una costanza nel numero cromosomico attraverso le generazioni di una particolare specie. Infatti se ciò non avvenisse la fusione di due cellule dipolidi produrrebbe uno zigote con il doppio dei cromosomi presenti nella generazione parentale, ad esempio nell'uomo il numero dei cromosomi in una cellula diploide è 46, la fusione di due cellule diploidi produrrebbe in seguito uno zigote con 92 cromosomi, nelle generazione successiva il doppio!
Inoltre errori nella segregazione dei cromosomi durante le fasi meiotiche possono portare a delle anomalie a livello genetico, come il processo definito di non disgiunzione nel quale i cromosomi non segregano correttamente portando alla formazione di gameti che presentano un numero maggiore di cromosomi in un gamete rispetto al normale e in un altro gamete essere assenti, oppure essere presenti in un numero deficitario rispetto al normale. La stragrande maggioranza di queste anomalie sono letali sia che coinvolgano cromosomi autosomici o sessuali, in alcuni casi come la trisomia 21 che causa la sindrome di down non è mortale ma porta modificazioni fenotipiche che possono essere molto debilitanti per l'individuo che ne è affetto.
In altri casi ancora a differenza dei casi in cui si verificano errori nella segregazione dei cromosomi, alcuni animali ibridi portano cromosomi non omologhi che non possono segregare correttamente. Un esempio è l'unione tra il cavallo e l'asino che produce il mulo che come molti sanno è sterile. Ciò è dovuto al fatto che il padre asino porta 30 cromosomi il cavallo 32 e molti di questi cromosomi sono troppo differenti per potersi appaiare, cio provoca formazione di gameti non vitali nel mulo con conseguente sterilità.
Abbiamo accennato al fatto che la meiosi svolge un ruolo di fondamentale importanza nell'assicurare la costanza del numero cromosomico nelle generazioni successive, ma allo stesso tempo svolge un ruolo primario nell'assicurare la variabilità genetica nelle generazioni. Infatti la meiosi presenta due aspetti di fondamentale importanza nell'assicurare la variabilità genetica. Primo è del tutto casuale quale dei cromosomi migri verso un determinato polo della cellula, 2) gameti diversi portano combinazioni genetiche differenti. Inoltre dobbiamo tenere conto anche del fatto che più aumenta l'assortimento di combinazioni geniche in una popolazione maggiori saranno le possibilità che alcuni individui possiedano combinazioni di alleli che possono permettere la sopravvivenza di un individuo in un particolare ambiente, e inoltra maggiore sarà il numero di cromosomi maggiore la variabilità genetica. Ad esempio negli esseri umani il numero di cromosomi in un gamete è 23 l'assortimento casuale di gameti con i rispettivi cromosomi potrebbe produrre 2 elevato alla 23 tipi di gameti differenti...(quindi 2 elevata alla 23 combinazioni genotipiche differenti).
Una seconda caratteristica poi è il mescolamento dei cromosomi che avviene durante il processo del crossing over in cui i cromosomi scambiano tra di loro segmenti di materiale genetico, contribuendo alla variabilità genetica, senza contare che la riproduzione sessuale amplifica questa variabilità, durante la riproduzione di uno qualsiasi dei tanti spermatozooi geneticamente diversi può fecondare uno dei tanti ovuli con la propria distinta identità genetica.
La meiosi è costituita da due cicli di divisione cellulare.
Caratteristica principale della meiosi è che a differenza della mitosi avengono due divisioni nucleari anzichè una sola.
Nella meiosi I l'evento principale è costituito dall'appaiamento dei cromosomi omologhi, dallo scambio di materiale genetico tra i cromosomi e infine dalla loro separazione.

Profase I: è la fase più lunga del processo meiotico, è il momento in cui avviene il crossing over.
La profase può essere suddivisa in vari sottostadi, il primo è noto come leptotene; all'avvio della meiosi I il dna è stato già duplicato, è la fsi in cui i cromosomi iniziano ad ispessirsi. Tutti i cromosomi sono già stati duplicati all'inizio della meiosi I ed ognuno è costituito da un cromatidio fratello attaccati d una struttura nota come centromero. In questo stadio i cromatidi sono strettamente associati tra di loro da apparire come un unica identità. Il successivo stadio è noto come zigotene, inizia questo stadio quando tutti i cromosomi omologhi sono appaiati tra di loro e inizian ad unirsi come una iusura lampo, a formare la sinapsi. QUesto processo avviene grazie all'azione e alla formazione di una elaborata e complessa struttura proteica nota come complesso sinaptinemale, è importante la formazione di tale complesso affinchè l'appaiamento dei cromosomi avvenga con estrema precisione, giustapponendo le stesse regioni genetiche dei due cromosomi.
Il pachitene; è lo stadio in cui avviene il crossing over, i cromosomi sono ora appaiati per tutta la loro lunghezza, i cromosomi X e Y sono differenti; l'appaiamento nel loro caso avviene solo in prossimità di particolari zone omolghe che permettono un appaiamento limitato. Durante il pachitene iniziano a formarsi delle particolari strutture denominate noduli di ricombinazione. A livello dei noduli di ricombinazione avviene uno scambio di materiale genomico tra i cromatidi non fratelli (materni e paterni). Lo scambio noto come crossing over, ha come risultato la ricombinazione di materiale genetico. Come risultato ogni cromosoma della coppia potrebbe non essere più di derivazione paterna e materna, ognuno comunque ha ancora un entità genomica completa dal momento che non vi è interruzione nell'informazione genetica.

Metafase I: la differenza tra il fuso che si viene a formare durante questo processo e quello che si forma nella metafase mitotica è che durante la meiosi ogni cromatidio fratello possiede un cinetocore in grado di poter interaginre con i microtubuli che originano dai poli della opposti al fuso.
Inoltre in meiosi I i cinetocori dei cromatidi fratelli vanno incontro a fusione, ciò farà si che ogni cromosoma contenga un solo cinetocore.
La presenza di un solo cinetocore ha una conseguenza, durante l'anafase non vi sono forze opposte in grado di tirare i cromatidi fratelli da un lato e dall'atro. Nella metafase I al contrario le strutture dei microtubulu provenienti da poli opposti si attaccano ai cinetocori dei cromosomi omologhi.
I cromosomi in questo modo sono allineati nella piastra metafasica, i cinetocori dei cromosoi materni e paterni si affaceranno ai poli opposti del fuso, posizionando i cromosomi omologhi in modo tale che durante l'anafase possano venire tirati in direzioni opposte.
Dal momento che l'allineamento e l'aggancio di ciascun bivalente alle fibre del fuso sono indipendenti da quelli di qualsiasi altro bivalente, i cromosomi che si affacceranno ad un polo saranno un miscuglio casuale di cromosomi di origine materna e paterna.
Anafase I: al momento dell'anafase I i chiasmi che uniscono i cromosomi omologhi si dissolvono, permettendo agli omologhi materni e paterni di muoversi verso i poli del fuso.
Nella prima divisione meiotica i centromeri non si dividono come in meiosi; così per ogni bivalente migregà verso ciascun polo solo un cromosoma, costituito da due cromatidi fratelli uniti al centromero.
Durante questa fase avviene la ricombinazione tramite il processo definito del crossing over, il quale riveste un ruolo di fondamentale importanza nella segregzione dei cromosomi omologhi durante la prima divisione meiotica.
Questo perchè i chiasmi, mantenendo uniti i cromosomi omologhi, assicurano anche che i centromeri rimangano attaccati ai poli opposti del fuso per tutta la durata della metafase.
Quando in un bivalente la ricombinazione non avviene si possono verificare dei grvi errori nell'aggncio, e nel trasporto dei cromosomi, che possono portare ad un movimento dei cromosomi omologhi verso lo stesso polo invece che tra i poli opposti.
Telofase I: Durante il processo della telofase iniziano a formarsi le membrane nucleari che si formano attorno ai cromosomi che sono migrati ai poli.
I nuclei figli che si stanno per formare contengono ciascuno metà dei cromosomi del nucleo parentale, ma ciascun cromosoma è costituito da cromatidi fratelli uniti al centromero, durante la meiosi il numero di cromosomi si riduce alla metà del normale numero diploide (quindi si ha in questo momento un corredo aploide).

Le fasi che caraterizzano la meiosi II sono più o meno identiche alla meiosi I:
Profase II: Nell'interfase precedente alla meiosi I i cromosomi si sono decondensati, ora si ricondensano al termine della profase II la membrana nucleare si dissolve nuovamente e si riforma il fuso.
Metafase II: durante tale fase i cinetocori dei cromatidi fratelli si attaccano al fibre del fuso, come nella metafase mitotica.
Tale fase però possiede una caratteristica fondamentale che la distingue da quella mitotica. Il numero dei cromosomi è la metà di quello presente nella metafase mitotica e nella maggior parte dei cromosomi i due cromatidi fratelli non sono più perfettamente identici a causa della ricombinazione avvenuta durante la meiosi I, i cromatidi possiedono ancora gli stessi geni ma possono portare combinazioni differenti di alleli.
Anafase II: come accade nella mitosi anche in questa fase i centromeri si separano e ciò permette ai cromatidi di migrare verso i poli opposti.
Telofase II: si sono formate quatttro cellule figlie si riformano le membrane e la citochinesi colloca i nuclei in cellula distinte. Il risultato è la formazione di quattro gameti aploidi .Alla fine di questa fase ognuno dei gameti ha lo stesso numero di cromosomi della cellula da cui si origina. La meiosi due per tale motivo viene definito divisione equazionale.
Iscriviti a:
Commenti (Atom)